Onychophora, velvet worms
Eoperipatus butleri
Eoperipatus butleri

Pulau Ubin : 13-4-24
Onychophora commonly known as velvet worms (due to their velvety texture and somewhat wormlike appearance) or more ambiguously as peripatus (after the first described genus, Peripatus), is a phylum of elongate, soft-bodied, many-legged animals.
In appearance they have variously been compared to worms with legs, caterpillars, and slugs. They prey upon other invertebrates, which they catch by ejecting an adhesive slime.
Approximately 200 species of velvet worms have been described, although the true number of species is likely greater.
The two extant families of velvet worms are Peripatidae and Peripatopsidae.
They show a peculiar distribution, with the peripatids being predominantly equatorial and tropical, while the peripatopsids are all found south of the equator.
It is the only phylum within Animalia that is wholly endemic to terrestrial environments, at least among extant members.
Velvet worms are generally considered close relatives of the Arthropoda and Tardigrada, with which they form the proposed taxon Panarthropoda. This makes them of palaeontological interest, as they can help reconstruct the ancestral arthropod.
Only two fossil species are confidently assigned to as onychophorans: Antennipatus from the Late Carboniferous, and Cretoperipatus from the Late Cretaceous, the latter belonging to Peripatidae.
In modern zoology, they are particularly renowned for their curious mating behaviours and the bearing of live young in some species.
Velvet worms are segmented animals with a flattened cylindrical body cross-section and rows of
unstructured body appendages known as oncopods or lobopods (informally: stub feet).
They reach lengths between 0.1 and 22 cm (0.04–8.66 in) depending on species, with the smallest known being Ooperipatellus nanus and the largest known is Mongeperipatus solorzanoi.
The number of leg pairs ranges from as few as 13 (in Ooperipatellus nanus) to as many as 43 (in Plicatoperipatus jamaicensis).
Their skin consists of numerous, fine transverse rings and is often inconspicuously coloured orange, red or brown, but sometimes also bright green, blue, gold or white, and occasionally patterned with other colours.
Segmentation is outwardly inconspicuous, and identifiable by the regular spacing of the pairs of legs and in the regular arrangement of skin pores, excretion organs and concentrations of nerve cells.
The individual body sections are largely unspecialised, even the head develops only a little differently from the abdominal segments.
Segmentation is apparently specified by the same gene as in other groups of animals, and is activated in each case, during embryonic development, at the rear border of each segment and in the growth zone of the stub feet.
Although onychophorans fall within the protostome group, their early development has a deuterostome trajectory, (with the mouth and anus forming separately) this trajectory is concealed by the rather sophisticated processes which occur in early development.
The stub feet that characterise the velvet worms are conical, baggy appendages of the body, which are internally hollow and have no joints.
Although the number of feet can vary considerably between species, their structure is basically very similar. Rigidity is provided by the hydrostatic pressure of their fluid contents, and movement is usually obtained passively by stretching and contraction of the animal's entire body.
However, each leg can also be shortened and bent by internal muscles. Due to the lack of joints, this bending can take place at any point along the sides of the leg.
In some species, two different organs are found within the feet :
* Crural glands are situated at the shoulder of the legs, extending into the body cavity. They open outwards at the crural papillae—small wart-like bumps on the belly side of the leg—and secrete chemical messenger materials called pheromones.
Their name comes from the Latin cruralis meaning "of the legs.
* Coxal vesicles are pouches located on the belly side of the leg, which can be everted and probably serve in water absorption.
They belong to the family Peripatidae and are named from coxa, the Latin word for "hip".
On each foot is a pair of retractable, hardened (sclerotised) chitin claws, which give the taxon its scientific name: Onychophora is derived from the Ancient Greek: ονυχής, onyches, "claws"; and φέρειν, pherein, "to carry".
At the base of the claws are three to six spiny
"cushions" on which the leg sits in its resting position and on which the animal walks over smooth substrates. The claws are used mainly to gain a firm foothold on uneven terrain.
Velvet worms have hydrostatic skeletons, it doesn't have hard exoskeletons like arthropods. Instead, their fluid-filled body cavities are covered in a thin skin and kept rigid by their pressurized internal liquids. They move by the alteration of fluid pressure in the limbs as they extend and contract along the body.
They have velvety, water-proof skin. Their entire bodies are covered with papillae, tiny protrusions with bristles sensitive to touch and smell. The papillae are made up of overlapping scales, which gives the velvet worm its velvety appearance. It also makes their skin water-repellant.
Velvet worms are vulnerable to dehydration. Like insects, velvet worms breathe through holes along their bodies called tracheae.
Unlike insects, velvet worms cannot close these holes to prevent water loss, so they easily dry out. For this reason, velvet worms spend most of their time hidden in moist areas in the soil, under rocks, and in rotting logs. They're most active at night and during rainy weather.
Velvet worms are ambush predators, they use slime as a weapon, hunting other small invertebrates by night.
To subdue their prey, they squirt a sticky, quick
hardening slime from a pair of glands on their heads. After the prey is ensnared, the velvet worm bites into it, injecting digestive saliva that helps liquefy the insides for easier snacking.
The slime is energetically costly to make, so velvet worms will often eat any excess slime they have produced to shore up their reserves.
Eoperipatus is a Southeast Asian genus of velvet worm in the family Peripatidae.
The number of legs in this genus varies within species as well as among species and ranges from 22 pairs (in E. butleri) to 25 pairs (in E. horsti and E. weldoni).
Females are usually larger than males and typically only produce young once in their lives.
Most velvet worms give birth to live young. Female velvet worms can store sperm for many months before using them to fertilize their eggs.
Their gestation period can last up to 15 months in some species. Most give birth to live young, although a few species lay eggs. Young velvet worms are born fully developed and looking like miniature versions of the adults.
They vary in how they mate and how young are born, some velvet worms lay eggs, and others produce live young, but the majority undergo a process known as ovoviviparity. In this process, embryos develop inside an egg, but the egg is kept inside the mother’s body until the young are ready to hatch.
This genus exhibits lecithotrophic ovoviviparity, that is, mothers in this genus retain yolky eggs in their uteri.
The genus contains the following described species:
• Eoperipatus butleri Evans, 1901
• Eoperipatus horsti Evans, 1901
• Eoperipatus totoro Oliveira et al., 2013
• Eoperipatus weldoni Evans, 1901
Eoperipatus sumatranus (Sedgwick, 1888) is considered a nomen dubium by Oliveira et al. 2012.
First recorded in Vietnam, Eoperipatus has now been found distributed throughout South-East Asia.
An undescribed species is known to occur in Thailand. In addition, unidentified onychophorans have also been observed in Borneo and in central Vietnam, north of the known distribution of Eoperipatus totoro. These may or may not represent distinct species. These species have been hard to study as they live in cryptic environments and have low population densities.
Eoperipatus butleri is a Malaysian species of velvet worm in the Peripatidae family.
Eoperipatus butleri was described by Richard Evans in 1901, from a single female specimen discovered by A. M. Butler in the Larut Hills of West Malaysia.
The generic name Eoperipatus is derived from an Ancient Greek combining form of ēṓs, meaning "dawn", and peripatos, meaning "walking about".
The specific name butleri is in honour of its discoverer. Although some have suggested that E. butleri is a junior synonym of E. weldoni, other authorities maintain that E. butleri is valid as a different species, citing the significant distance (over 300 km) between the type localities of these two species.
The dorsal surface is dark brown with pale spots and a darker brown mid-dorsal line extending from the first pair of oncopods (legs) to the anus.
The ventral surface is slightly lighter brown than the dorsal surface. The type specimen was 52 mm in length and 6 mm in width. The female of this species has 24 pairs of legs, but the male has only 22 leg pairs.
Phylum : Onychophora
Family : Peripatidae
Genus : Eoperipatus
Species : E. butleri
Binomial name Eoperipatus butleri
Evans, 1901
In appearance they have variously been compared to worms with legs, caterpillars, and slugs. They prey upon other invertebrates, which they catch by ejecting an adhesive slime.
Approximately 200 species of velvet worms have been described, although the true number of species is likely greater.
The two extant families of velvet worms are Peripatidae and Peripatopsidae.
They show a peculiar distribution, with the peripatids being predominantly equatorial and tropical, while the peripatopsids are all found south of the equator.
It is the only phylum within Animalia that is wholly endemic to terrestrial environments, at least among extant members.
Velvet worms are generally considered close relatives of the Arthropoda and Tardigrada, with which they form the proposed taxon Panarthropoda. This makes them of palaeontological interest, as they can help reconstruct the ancestral arthropod.
Only two fossil species are confidently assigned to as onychophorans: Antennipatus from the Late Carboniferous, and Cretoperipatus from the Late Cretaceous, the latter belonging to Peripatidae.
In modern zoology, they are particularly renowned for their curious mating behaviours and the bearing of live young in some species.
Velvet worms are segmented animals with a flattened cylindrical body cross-section and rows of
unstructured body appendages known as oncopods or lobopods (informally: stub feet).
They reach lengths between 0.1 and 22 cm (0.04–8.66 in) depending on species, with the smallest known being Ooperipatellus nanus and the largest known is Mongeperipatus solorzanoi.
The number of leg pairs ranges from as few as 13 (in Ooperipatellus nanus) to as many as 43 (in Plicatoperipatus jamaicensis).
Their skin consists of numerous, fine transverse rings and is often inconspicuously coloured orange, red or brown, but sometimes also bright green, blue, gold or white, and occasionally patterned with other colours.
Segmentation is outwardly inconspicuous, and identifiable by the regular spacing of the pairs of legs and in the regular arrangement of skin pores, excretion organs and concentrations of nerve cells.
The individual body sections are largely unspecialised, even the head develops only a little differently from the abdominal segments.
Segmentation is apparently specified by the same gene as in other groups of animals, and is activated in each case, during embryonic development, at the rear border of each segment and in the growth zone of the stub feet.
Although onychophorans fall within the protostome group, their early development has a deuterostome trajectory, (with the mouth and anus forming separately) this trajectory is concealed by the rather sophisticated processes which occur in early development.
The stub feet that characterise the velvet worms are conical, baggy appendages of the body, which are internally hollow and have no joints.
Although the number of feet can vary considerably between species, their structure is basically very similar. Rigidity is provided by the hydrostatic pressure of their fluid contents, and movement is usually obtained passively by stretching and contraction of the animal's entire body.
However, each leg can also be shortened and bent by internal muscles. Due to the lack of joints, this bending can take place at any point along the sides of the leg.
In some species, two different organs are found within the feet :
* Crural glands are situated at the shoulder of the legs, extending into the body cavity. They open outwards at the crural papillae—small wart-like bumps on the belly side of the leg—and secrete chemical messenger materials called pheromones.
Their name comes from the Latin cruralis meaning "of the legs.
* Coxal vesicles are pouches located on the belly side of the leg, which can be everted and probably serve in water absorption.
They belong to the family Peripatidae and are named from coxa, the Latin word for "hip".
On each foot is a pair of retractable, hardened (sclerotised) chitin claws, which give the taxon its scientific name: Onychophora is derived from the Ancient Greek: ονυχής, onyches, "claws"; and φέρειν, pherein, "to carry".
At the base of the claws are three to six spiny
"cushions" on which the leg sits in its resting position and on which the animal walks over smooth substrates. The claws are used mainly to gain a firm foothold on uneven terrain.
Velvet worms have hydrostatic skeletons, it doesn't have hard exoskeletons like arthropods. Instead, their fluid-filled body cavities are covered in a thin skin and kept rigid by their pressurized internal liquids. They move by the alteration of fluid pressure in the limbs as they extend and contract along the body.
They have velvety, water-proof skin. Their entire bodies are covered with papillae, tiny protrusions with bristles sensitive to touch and smell. The papillae are made up of overlapping scales, which gives the velvet worm its velvety appearance. It also makes their skin water-repellant.
Velvet worms are vulnerable to dehydration. Like insects, velvet worms breathe through holes along their bodies called tracheae.
Unlike insects, velvet worms cannot close these holes to prevent water loss, so they easily dry out. For this reason, velvet worms spend most of their time hidden in moist areas in the soil, under rocks, and in rotting logs. They're most active at night and during rainy weather.
Velvet worms are ambush predators, they use slime as a weapon, hunting other small invertebrates by night.
To subdue their prey, they squirt a sticky, quick
hardening slime from a pair of glands on their heads. After the prey is ensnared, the velvet worm bites into it, injecting digestive saliva that helps liquefy the insides for easier snacking.
The slime is energetically costly to make, so velvet worms will often eat any excess slime they have produced to shore up their reserves.
Eoperipatus is a Southeast Asian genus of velvet worm in the family Peripatidae.
The number of legs in this genus varies within species as well as among species and ranges from 22 pairs (in E. butleri) to 25 pairs (in E. horsti and E. weldoni).
Females are usually larger than males and typically only produce young once in their lives.
Most velvet worms give birth to live young. Female velvet worms can store sperm for many months before using them to fertilize their eggs.
Their gestation period can last up to 15 months in some species. Most give birth to live young, although a few species lay eggs. Young velvet worms are born fully developed and looking like miniature versions of the adults.
They vary in how they mate and how young are born, some velvet worms lay eggs, and others produce live young, but the majority undergo a process known as ovoviviparity. In this process, embryos develop inside an egg, but the egg is kept inside the mother’s body until the young are ready to hatch.
This genus exhibits lecithotrophic ovoviviparity, that is, mothers in this genus retain yolky eggs in their uteri.
The genus contains the following described species:
• Eoperipatus butleri Evans, 1901
• Eoperipatus horsti Evans, 1901
• Eoperipatus totoro Oliveira et al., 2013
• Eoperipatus weldoni Evans, 1901
Eoperipatus sumatranus (Sedgwick, 1888) is considered a nomen dubium by Oliveira et al. 2012.
First recorded in Vietnam, Eoperipatus has now been found distributed throughout South-East Asia.
An undescribed species is known to occur in Thailand. In addition, unidentified onychophorans have also been observed in Borneo and in central Vietnam, north of the known distribution of Eoperipatus totoro. These may or may not represent distinct species. These species have been hard to study as they live in cryptic environments and have low population densities.
Eoperipatus butleri is a Malaysian species of velvet worm in the Peripatidae family.
Eoperipatus butleri was described by Richard Evans in 1901, from a single female specimen discovered by A. M. Butler in the Larut Hills of West Malaysia.
The generic name Eoperipatus is derived from an Ancient Greek combining form of ēṓs, meaning "dawn", and peripatos, meaning "walking about".
The specific name butleri is in honour of its discoverer. Although some have suggested that E. butleri is a junior synonym of E. weldoni, other authorities maintain that E. butleri is valid as a different species, citing the significant distance (over 300 km) between the type localities of these two species.
The dorsal surface is dark brown with pale spots and a darker brown mid-dorsal line extending from the first pair of oncopods (legs) to the anus.
The ventral surface is slightly lighter brown than the dorsal surface. The type specimen was 52 mm in length and 6 mm in width. The female of this species has 24 pairs of legs, but the male has only 22 leg pairs.
Phylum : Onychophora
Family : Peripatidae
Genus : Eoperipatus
Species : E. butleri
Binomial name Eoperipatus butleri
Evans, 1901


Bipalium graffi
Broadhead Planarian
Broadhead Planarian
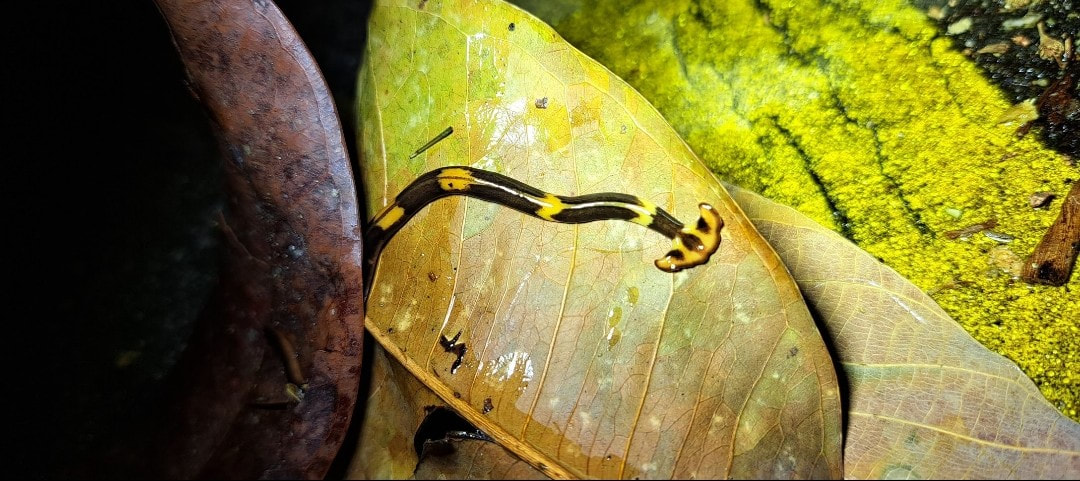
Bipaliinae is a subfamily of land planarians found mainly in Madagascar, the Indian subcontinent and Southeast Asia, although some species have been introduced worldwide.
The subfamily Bipaliinae is characterized by having a semilunar head that gives them the common name "hammerhead worms". The head has peripheral sensory pits on the ventral side and small peripheral eyes on the dorsal side.
Species in the subfamily Bipaliinae are grouped in the following genera:
• Bipalium Stimpson, 1857
• Humbertium Ogren & Sluys, 2001
• Novibipalium Kawakatsu et al., 1998
• Diversibipalium Kawakatsu et al., 2002
Several hammerhead flatworms have become invasive, the most famous being Bipalium kewense, now in many countries in all continents except the Antarctica.
A 2022 study used occurrence records from online databases, including iNaturalist, and climatic and soil variable to model the potential distribution of five species of hammerhead flatworms, namely B. kewense, B. adventitium, B. pennsylvaanicum, B. vagum, and Diversibipalium multilineatum.
It was found that the five species could invade Southeast Asia, New Zealand, Eastern Australian, a part of South America, eastern USA, western Europe and central Africa.
The name Bipalium comes from Latin bi-, "two" + pala, "shovel" or "spade", because species in this genus resemble a pickaxe.
The genus Bipalium was initially defined by Stimpson to include land planarians with the head broadened, forming a head plate. Later, in 1899, Ludwig von Graff divided it into three genera according to the shape of the head:
• Bipalium: with a well-developed head plate, much broader than long, and with elongated lateral auricles
• Perocephalus: rudimentary head plate, not much broader than the body
• Placocephalus: flat head plate with a circular outline
Josef Müller, in 1902, considered that no sufficient anatomical basis existed for this distinction, and reunited them under Bipalium. Later, von Graff accepted Müller's conclusions.
Towards the end of the 20th century, Robert E. Ogren and Masaharu Kawakatsu started a series of
publications called "The Land Planarian Index series" in which they reviewed and organized all taxonomic information regarding land planarians.
At first, they retained all broad-headed planarians in the genus Bipalium, but later split them into four
genera based on the anatomy of the reproductive organs: Bipalium, Novibipalium, Humbertium, and Diversibipalium.
Under this scenario, the genus Bipalium is defined as containing broad-headed land planarians with simple copulatory organs, without accessory ducts or copulatory bursa, and with a fold of tissue separating the male and female exit ducts.
To feed on their prey, species of Bipalium often evert their pharynges from their mouths located on the
midventral portion of their bodies, and secrete enzymes that begin the digestion of the prey. The liquefied tissues are sucked into the branching gut of the flatworms by ciliary action.
(The pharynx (plural: pharynges) is the part of the throat behind the mouth and nasal cavity, and above
the esophagus and trachea (the tubes going down
to the stomach and the lungs). It is found in vertebrates and invertebrates, though its structure varies across species.
The pharynx carries food and air to the esophagus and larynx respectively. The flap of cartilage called
the epiglottis stops food from entering the larynx)
Reproduction in Bipalium may be asexual or sexual and all species are hermaphroditic
In reproductive biology, a hermaphrodite (/hərˈmæfrəˌdaɪt/) is an organism that has both kinds of reproductive organs and can produce both gametes associated with male and female sexes.
Bipalium graffi is a species of flatworms in the family land planarians. They rely on ciliary gliding to move around. Ciliary, a type of locomotion in which an animal moves on a secreted layer of mucus, propelled by the beating of cilia. It is characteristic of small, soft-bodied invertebrates.
Phylum : Platyhelminthes
Order : Tricladida
Family : Geoplanidae
Subfamily : Bipaliinae
Genus : Bipalium
Stimpson, 1857
Type species Bipalium fuscatum
Stimpson, 1857
The subfamily Bipaliinae is characterized by having a semilunar head that gives them the common name "hammerhead worms". The head has peripheral sensory pits on the ventral side and small peripheral eyes on the dorsal side.
Species in the subfamily Bipaliinae are grouped in the following genera:
• Bipalium Stimpson, 1857
• Humbertium Ogren & Sluys, 2001
• Novibipalium Kawakatsu et al., 1998
• Diversibipalium Kawakatsu et al., 2002
Several hammerhead flatworms have become invasive, the most famous being Bipalium kewense, now in many countries in all continents except the Antarctica.
A 2022 study used occurrence records from online databases, including iNaturalist, and climatic and soil variable to model the potential distribution of five species of hammerhead flatworms, namely B. kewense, B. adventitium, B. pennsylvaanicum, B. vagum, and Diversibipalium multilineatum.
It was found that the five species could invade Southeast Asia, New Zealand, Eastern Australian, a part of South America, eastern USA, western Europe and central Africa.
The name Bipalium comes from Latin bi-, "two" + pala, "shovel" or "spade", because species in this genus resemble a pickaxe.
The genus Bipalium was initially defined by Stimpson to include land planarians with the head broadened, forming a head plate. Later, in 1899, Ludwig von Graff divided it into three genera according to the shape of the head:
• Bipalium: with a well-developed head plate, much broader than long, and with elongated lateral auricles
• Perocephalus: rudimentary head plate, not much broader than the body
• Placocephalus: flat head plate with a circular outline
Josef Müller, in 1902, considered that no sufficient anatomical basis existed for this distinction, and reunited them under Bipalium. Later, von Graff accepted Müller's conclusions.
Towards the end of the 20th century, Robert E. Ogren and Masaharu Kawakatsu started a series of
publications called "The Land Planarian Index series" in which they reviewed and organized all taxonomic information regarding land planarians.
At first, they retained all broad-headed planarians in the genus Bipalium, but later split them into four
genera based on the anatomy of the reproductive organs: Bipalium, Novibipalium, Humbertium, and Diversibipalium.
Under this scenario, the genus Bipalium is defined as containing broad-headed land planarians with simple copulatory organs, without accessory ducts or copulatory bursa, and with a fold of tissue separating the male and female exit ducts.
To feed on their prey, species of Bipalium often evert their pharynges from their mouths located on the
midventral portion of their bodies, and secrete enzymes that begin the digestion of the prey. The liquefied tissues are sucked into the branching gut of the flatworms by ciliary action.
(The pharynx (plural: pharynges) is the part of the throat behind the mouth and nasal cavity, and above
the esophagus and trachea (the tubes going down
to the stomach and the lungs). It is found in vertebrates and invertebrates, though its structure varies across species.
The pharynx carries food and air to the esophagus and larynx respectively. The flap of cartilage called
the epiglottis stops food from entering the larynx)
Reproduction in Bipalium may be asexual or sexual and all species are hermaphroditic
In reproductive biology, a hermaphrodite (/hərˈmæfrəˌdaɪt/) is an organism that has both kinds of reproductive organs and can produce both gametes associated with male and female sexes.
Bipalium graffi is a species of flatworms in the family land planarians. They rely on ciliary gliding to move around. Ciliary, a type of locomotion in which an animal moves on a secreted layer of mucus, propelled by the beating of cilia. It is characteristic of small, soft-bodied invertebrates.
Phylum : Platyhelminthes
Order : Tricladida
Family : Geoplanidae
Subfamily : Bipaliinae
Genus : Bipalium
Stimpson, 1857
Type species Bipalium fuscatum
Stimpson, 1857





Leptogoniulus sorornus
Giant millipede
Giant millipede

Spirostreptida is an order of long, cylindrical millipedes. There are approximately 1000 described species, making Spirostreptida the third largest order of millipedes after Polydesmida and Chordeumatida.
Spirostreptida are generally large, long and cylindrical, with 30 to 90 body rings. Eyes are present in most. Spirostreptida contains mainly tropical species.
Like most millipede groups, they have a fragmentary fossil record. The oldest record of the group is the extinct family Electrocambalidae, which is known from the Burmese amber of Myanmar, dating to the Cenomanian stage of the Late Cretaceous around 99 million years ago, which belongs to the suborder Cambalidea.
Harpagophoridae is a family of round-backed millipedes of the order Spirostreptida. The family includes 269 species belonging to 55 genera, distributed in Africa, Southeast Asia, and the East Indies, as well as a few Indian Oceanic islands. Two subfamilies are recognized.
This white-legged giant millipede of the family Harpagophoridae emits utra violet lights and measured approximately 7cm to 8 cm long was photographed at Mandai T7 along the side road of the Nature Reserve.
Phylum: Arthropoda
Subphylum: Myriapoda
Class: Diplopoda
Order: Spirostreptida
Family: Harpagophoridae
Attems, 1909
Gene: Leptogoniulus
Species : Leptogoniulus sorornus (Butler, 1876).
Spirostreptida are generally large, long and cylindrical, with 30 to 90 body rings. Eyes are present in most. Spirostreptida contains mainly tropical species.
Like most millipede groups, they have a fragmentary fossil record. The oldest record of the group is the extinct family Electrocambalidae, which is known from the Burmese amber of Myanmar, dating to the Cenomanian stage of the Late Cretaceous around 99 million years ago, which belongs to the suborder Cambalidea.
Harpagophoridae is a family of round-backed millipedes of the order Spirostreptida. The family includes 269 species belonging to 55 genera, distributed in Africa, Southeast Asia, and the East Indies, as well as a few Indian Oceanic islands. Two subfamilies are recognized.
This white-legged giant millipede of the family Harpagophoridae emits utra violet lights and measured approximately 7cm to 8 cm long was photographed at Mandai T7 along the side road of the Nature Reserve.
Phylum: Arthropoda
Subphylum: Myriapoda
Class: Diplopoda
Order: Spirostreptida
Family: Harpagophoridae
Attems, 1909
Gene: Leptogoniulus
Species : Leptogoniulus sorornus (Butler, 1876).





Bipalium vagum
Hammerhead worm
Hammerhead worm

Bipalium vagum is a land planarian in the subfamily Bipaliinae. It has been accidentally introduced in the United States, Bermuda and various islands in the Caribbean.
Bipalium vagum is a relatively small species of Bipalium, measuring about 25 mm (0.98 in) in length. The head varies from entirely black to dark brown with two black patches separated by a lighter ground color. The neck has a black collar interrupted only at the creeping sole. The dorsal color of the body is light brown and a broad black stripe runs longitudinally from the black collar to the posterior end. Laterally, there are two diffuse dark brown stripes.
Differently from other invasive species of Bipalium, which feed on earthworms, B. vagum seems to feed exclusively on gastropods. As it seems to be spreading throughout the Caribbean and southern United States, there is certain concern on its possible impacts on the native gastropod fauna.
This species Bipaluim vagum was found in old upper thomson road on 5th Aug 2021 during nite photography and likely this individuals were introduced possibly with imported ornamental plants.
Phylum: Platyhelminthes
Class: Rhabditophora
Order: Tricladida
Suborder: Continenticola
Family: Geoplanidae
Subfamily: Bipaliinae
Genus: Bipalium
Species: B. vagum
Binomial name Bipalium vagum
Jones & Sterrer, 2005
Bipalium vagum is a relatively small species of Bipalium, measuring about 25 mm (0.98 in) in length. The head varies from entirely black to dark brown with two black patches separated by a lighter ground color. The neck has a black collar interrupted only at the creeping sole. The dorsal color of the body is light brown and a broad black stripe runs longitudinally from the black collar to the posterior end. Laterally, there are two diffuse dark brown stripes.
Differently from other invasive species of Bipalium, which feed on earthworms, B. vagum seems to feed exclusively on gastropods. As it seems to be spreading throughout the Caribbean and southern United States, there is certain concern on its possible impacts on the native gastropod fauna.
This species Bipaluim vagum was found in old upper thomson road on 5th Aug 2021 during nite photography and likely this individuals were introduced possibly with imported ornamental plants.
Phylum: Platyhelminthes
Class: Rhabditophora
Order: Tricladida
Suborder: Continenticola
Family: Geoplanidae
Subfamily: Bipaliinae
Genus: Bipalium
Species: B. vagum
Binomial name Bipalium vagum
Jones & Sterrer, 2005

Bipalium rauchi
Hammerhead worms
Hammerhead worms

Diversibipalium rauchi , Synonym (Bipalium rauchi)
Bipalium is a genus of large predatory land planarians. They are often loosely called "hammerhead worms" or "broadhead planarians" because of the distinctive shape of their head region.
Land planarians are unique in that they possess a "creeping sole", a highly ciliated region on the ventral
epidermis that helps them to creep over the substrate.
Several species are considered as invasive to the United States and to Europe. Some studies have begun the investigation of the evolutionary ecology of these invasive planarians.
The genus Bipalium was initially defined by Stimpson to include land planarians with the head broadened, forming a head plate. Later, in 1899, Ludwig von Graff divided it into three genera according to the shape of the head.
Bipalium: with a well-developed head plate, much broader than long, and with elongated lateral auricles.
Perocephalus: rudimentary head plate, not much broader than the body.
Placocephalus: flat head plate with a circular outline.
Josef Müller, in 1902, considered that no sufficient anatomical basis existed for this distinction, and reunited them under Bipalium. Later, von Graff accepted Müller's conclusions.
Towards the end of the 20th century, Robert E. Ogren and Masaharu Kawakatsu started a series of publications called "The Land Planarian Index series" in which they reviewed and organized all taxonomic information regarding land planarians.
At first, they retained all broad-headed planarians in the genus Bipalium, but later split them into four genera based on the anatomy of the reproductive organs: Bipalium, Novibipalium, Humbertium, and Diversibipalium.
Under this scenario, the genus Bipalium is defined as containing broad-headed land planarians with simple copulatory organs, without accessory ducts or copulatory bursa, and with a fold of tissue separating the male and female exit ducts.
The name Bipalium comes from Latin bi-, “two” + pala, “shovel” or “spade”, because species in this genus resemble a pickaxe.
Bipalium species are predatory. Some species prey on earthworms, while others may also feed on mollusks. These flatworms can track their prey. When captured, earthworms begin to react to the attack, but a flatworm uses the muscles in its body, as well as sticky secretions, to attach itself to the earthworm to prevent escape.
The planarians cover, or cap, the prostomium,
peristomium, and anterior end to stop the violent reaction by the earthworm.
To feed on their prey, species of Bipalium often evert their pharynges from their mouths, located on the midventral portion of their bodies, and secrete enzymes that begin the digestion of the prey. The liquefied tissues are sucked into the branching gut of the flatworms by ciliary action.
Reproduction in Bipalium may be asexual or sexual and all species are hermaphroditic.
B. adventitium reproduces sexually and creates egg capsules, which hatch around 3 weeks post-deposition. The egg capsules have a tough exterior and generally contain multiple juveniles.
B. kewense have rarely been observed using egg capsules as a primary method of reproduction. Asexual fragmentation is its main reproductive strategy in temperate regions and most individuals never develop sexual organs. Juveniles of this species, unlike B. adventitium, do not appear the same coloration as parents in their early days.
Recently, the presence of tetrodotoxin, a very potent neurotoxin, was recorded in B. adventitium and B. kewense. It is the first record of tetrodotoxin in terrestrial invertebrates.
Little is known about the ecology of terrestrial planarians, but research has been done on different genera and species, including several native and invasive species in Brazil, Arthurdendyus triangulatus, Rhynchodemus and Bipalium.
Other animals rarely devour land planarians, since surface secretions appear distasteful, if not toxic. Protozoans, including flagellates, ciliates, sporozoans, and nematodes have been detected in land planarians. Because of their cannibalistic habit, land planarians may be their own worst enemy.
Specimens of Bipalium adventitium are characterized by a single dark dorsal stripe. They were first discovered in the US in California and New York but have been found in Illinois as well as most northern states.
Bipalium kewense have five dark dorsal stripes and a partial dark collar. They have undergone several name changes since their discovery in North America.
B. kewense is commonly found across the southern regions of the US. B. kewense is found worldwide in tropical and subtropical regions.
Bipalium pennsylvanicum is characterized by its dark brown head and three dorsal stripes. Thus far, it has only been found in Pennsylvania and in coastal South Carolina in 2014.
Bipalium vagum is characterized by two dark dorsal blotches on the head, a thick black band around the neck, and three dark dorsal stripes. This species has recently been found in several Gulf Coast states.
Recently it was found in Mumbai, India
Phylum: Platyhelminthes
Class: Rhabditophora
Order: Tricladida
Suborder: Continenticola
Family: Geoplanidae
Subfamily: Bipaliinae
Genus: Bipalium
Stimpson, 1857
Land planarians are unique in that they possess a "creeping sole", a highly ciliated region on the ventral
epidermis that helps them to creep over the substrate.
Several species are considered as invasive to the United States and to Europe. Some studies have begun the investigation of the evolutionary ecology of these invasive planarians.
The genus Bipalium was initially defined by Stimpson to include land planarians with the head broadened, forming a head plate. Later, in 1899, Ludwig von Graff divided it into three genera according to the shape of the head.
Bipalium: with a well-developed head plate, much broader than long, and with elongated lateral auricles.
Perocephalus: rudimentary head plate, not much broader than the body.
Placocephalus: flat head plate with a circular outline.
Josef Müller, in 1902, considered that no sufficient anatomical basis existed for this distinction, and reunited them under Bipalium. Later, von Graff accepted Müller's conclusions.
Towards the end of the 20th century, Robert E. Ogren and Masaharu Kawakatsu started a series of publications called "The Land Planarian Index series" in which they reviewed and organized all taxonomic information regarding land planarians.
At first, they retained all broad-headed planarians in the genus Bipalium, but later split them into four genera based on the anatomy of the reproductive organs: Bipalium, Novibipalium, Humbertium, and Diversibipalium.
Under this scenario, the genus Bipalium is defined as containing broad-headed land planarians with simple copulatory organs, without accessory ducts or copulatory bursa, and with a fold of tissue separating the male and female exit ducts.
The name Bipalium comes from Latin bi-, “two” + pala, “shovel” or “spade”, because species in this genus resemble a pickaxe.
Bipalium species are predatory. Some species prey on earthworms, while others may also feed on mollusks. These flatworms can track their prey. When captured, earthworms begin to react to the attack, but a flatworm uses the muscles in its body, as well as sticky secretions, to attach itself to the earthworm to prevent escape.
The planarians cover, or cap, the prostomium,
peristomium, and anterior end to stop the violent reaction by the earthworm.
To feed on their prey, species of Bipalium often evert their pharynges from their mouths, located on the midventral portion of their bodies, and secrete enzymes that begin the digestion of the prey. The liquefied tissues are sucked into the branching gut of the flatworms by ciliary action.
Reproduction in Bipalium may be asexual or sexual and all species are hermaphroditic.
B. adventitium reproduces sexually and creates egg capsules, which hatch around 3 weeks post-deposition. The egg capsules have a tough exterior and generally contain multiple juveniles.
B. kewense have rarely been observed using egg capsules as a primary method of reproduction. Asexual fragmentation is its main reproductive strategy in temperate regions and most individuals never develop sexual organs. Juveniles of this species, unlike B. adventitium, do not appear the same coloration as parents in their early days.
Recently, the presence of tetrodotoxin, a very potent neurotoxin, was recorded in B. adventitium and B. kewense. It is the first record of tetrodotoxin in terrestrial invertebrates.
Little is known about the ecology of terrestrial planarians, but research has been done on different genera and species, including several native and invasive species in Brazil, Arthurdendyus triangulatus, Rhynchodemus and Bipalium.
Other animals rarely devour land planarians, since surface secretions appear distasteful, if not toxic. Protozoans, including flagellates, ciliates, sporozoans, and nematodes have been detected in land planarians. Because of their cannibalistic habit, land planarians may be their own worst enemy.
Specimens of Bipalium adventitium are characterized by a single dark dorsal stripe. They were first discovered in the US in California and New York but have been found in Illinois as well as most northern states.
Bipalium kewense have five dark dorsal stripes and a partial dark collar. They have undergone several name changes since their discovery in North America.
B. kewense is commonly found across the southern regions of the US. B. kewense is found worldwide in tropical and subtropical regions.
Bipalium pennsylvanicum is characterized by its dark brown head and three dorsal stripes. Thus far, it has only been found in Pennsylvania and in coastal South Carolina in 2014.
Bipalium vagum is characterized by two dark dorsal blotches on the head, a thick black band around the neck, and three dark dorsal stripes. This species has recently been found in several Gulf Coast states.
Recently it was found in Mumbai, India
Phylum: Platyhelminthes
Class: Rhabditophora
Order: Tricladida
Suborder: Continenticola
Family: Geoplanidae
Subfamily: Bipaliinae
Genus: Bipalium
Stimpson, 1857
Bipalium vagum, likely this individuals species were introduced possibly with imported ornamental plants.

Bipalium kewense


Bipalium kewense




Bipalium graffi
Helicorthomorpha holstii

Millipedes are a group of arthropods that are characterised by having two pairs of jointed legs on most body segments; they are known scientifically as the class Diplopoda, the name being derived from this feature.
Each double-legged segment is a result of two single segments fused together. Most millipedes have very elongated cylindrical or flattened bodies with more than 20 segments, while pill millipedes are shorter and can roll into a ball. Although the name "millipede" derives from the Latin for "thousand feet", no known species has 1,000; the record of 750 legs belongs to Illacme plenipes.
There are approximately 12,000 named species
classified into 16 orders and around 140 families, making Diplopoda the largest class of myriapods, an arthropod group which also includes centipedes and other multi-legged creatures.
Millipedes come in a variety of body shapes and sizes, ranging from 2 mm (0.08 in) to around 35 cm (14 in) in length and can have as few as eleven to over a hundred segments. They are generally black or brown in colour, although there are a few brightly coloured species, and some have aposematic colouring to warn that they are toxic, species of Motyxia produce cyanide as a chemical defence and are bioluminescent.
Body styles vary greatly between major millipede groups. In the basal subclass Penicillata, consisting of the tiny bristle millipedes, the exoskeleton is soft and uncalcified, and is covered in prominent setae or bristles.
All other millipedes, belonging to the subclass Chilognatha, have a hardened exoskeleton. The chilognaths are in turn divided into two infraclasses: the Pentazonia, containing relatively short-bodied groups such as pill millipedes, and the Helminthomorpha ("worm-like" millipedes), which contains the vast majority of species, with long, many-segmented bodies.
The head of a millipede is typically rounded above and flattened below and bears a pair of large mandibles in front of a plate-like structure called a gnathochilarium ("jaw lip").
The head contains a single pair of antennae with seven or eight segments and a group of sensory cones at the tip. Many orders also possess a pair of sensory organs known as the Tömösváry organs, shaped as small oval rings posterior and lateral to the base of the antennae. Their function is unknown, but they also occur in some centipedes, and are possibly used to measure humidity or light levels in the surrounding environment.
Millipede eyes consist of several simple flat lensed
nocelli arranged in a group or patch on each side of the head. These patches are also called ocular fields or ocellaria. Many species of millipedes, including the entire orders Polydesmida, Siphoniulida,
Glomeridesmida, Siphonophorida and Platydesmida, and cave-dwelling millipedes such as Causeyellan and Trichopetalum, had ancestors that could see but have subsequently lost their eyes and are blind.
Millipede bodies may be flattened or cylindrical, and are composed of numerous metameric segments, each with an exoskeleton consisting of four chitinous plates: a single plate above (the tergite), one at each side (pleurites), and a plate on the underside (sternite) where the legs attach.
In many millipedes, such as Merocheta and Juliformia, these plates are fused to varying degrees, sometimes forming a single cylindrical ring. The plates are typically hard, being impregnated with calcium salts. Because they can't close their permanently open spiracles and most species lack a waxy cuticle, millipedes are susceptible to water loss and with a few exceptions must spend most of their time in moist or humid environments.
The first segment behind the head is legless and known as a collum (from the Latin for neck or collar).
The second, third, and fourth body segments bear a single pair of legs each and are known as "haplosegments" (the three haplosegments are sometimes referred to as a "thorax").
The remaining segments, from the fifth to the posterior, are properly known as diplosegments or double segments, formed by the fusion of two embryonic segments. Each diplosegment bears two pairs of legs, rather than just one as in centipedes.
In some millipedes, the last few segments may be legless. The terms "segment" or "body ring" are often used interchangeably to refer to both haplo- and diplosegments. The final segment is known as the telson and consists of a legless preanal ring, a pair of anal valves (closeable plates around the anus), and a small scale below the anus.
Millipedes in several orders have keel-like extensions of the body-wall known as paranota, which can vary widely in shape, size, and texture; modifications include lobes, papillae, ridges, crests, spines and notches. Paranota may allow millipedes to wedge more securely into crevices, protect the legs, or make the millipede more difficult for predators to swallow.
The legs are composed of seven segments, and attach on the underside of the body. The legs of an individual are generally rather similar to each other, although often longer in males than females, and males of some species may have a reduced or enlarged first pair of legs.
The most conspicuous leg modifications are involved in reproduction, discussed below. Despite the common name, no millipede has been discovered with 1,000 legs: common species have between 34 and 400 legs, and the record is held by Illacme plenipes, with individuals possessing up to 750 legs – more than any other creature on Earth.
Millipedes breathe through two pairs of spiracles located ventrally on each segment near the base of the legs. Each opens into an internal pouch, and connects to a system of tracheae. The heart runs the entire length of the body, with an aorta stretching into the head. The excretory organs are two pairs of malpighian tubules, located near the mid-part of the gut. The digestive tract is a simple tube with two pairs of salivary glands to help digest the food.
Millipedes show a diversity of mating styles and structures. In the basal order Polyxenida (bristle millipedes), mating is indirect: males deposit spermatophores onto webs they secrete with special glands, and the spermatophores are subsequently picked up by females.
In all other millipede groups, males possess one or two pairs of modified legs called gonopods which are used to transfer sperm to the female during copulation.
The location of the gonopods differs between groups: in males of the Pentazonia they are located at the rear of the body and known as telopods and may also function in grasping females, while in the Helminthomorpha – the vast majority of species – they are located on the seventh body segment. A few species are parthenogenetic, having few, if any, males.
Gonopods occur in a diversity of shapes and sizes, and in the range from closely resembling walking legs to complex structures quite unlike legs at all.
In some groups, the gonopods are kept retracted within the body; in others they project forward parallel to the body. Gonopod morphology is the predominant means of determining species among millipedes: the structures may differ greatly between closely related species but very little within a species. The gonopods develop gradually from walking legs through successive moults until reproductive maturity.
The genital openings (gonopores) of both sexes are located on the underside of the third body segment (near the second pair of legs) and may be accompanied in the male by one or two penes which deposit the sperm packets onto the gonopods.
In the female, the genital pores open into paired small sacs called cyphopods or vulvae, which are covered by small hood-like lids, and are used to store the sperm after copulation. The cyphopod morphology can also be used to identify species. Millipede sperm lack flagella, a unique trait among myriapods.
In all except the bristle millipedes, copulation occurs with the two individuals facing one another.
Copulation may be preceded by male behaviours such as tapping with antennae, running along the back of the female, offering edible glandular secretions, or in the case of some pill-millipedes, stridulation or "chirping". During copulation in most millipedes, the male positions his seventh segment in front of the female's third segment, and may insert his gonopods to extrude the vulvae before bending his body to deposit sperm onto his gonopods and reinserting the "charged" gonopods into the female.
Females lay from ten to three hundred eggs at a time, depending on species, fertilising them with the stored sperm as they do so. Many species deposit the eggs on moist soil or organic detritus, but some construct nests lined with dried faeces, and may protect the eggs within silk cocoons. In most species, the female abandons the eggs after they are laid, but some species in the orders Platydesmida and Stemmiulida provide parental care for eggs and young.
The young hatch after a few weeks, and typically have only three pairs of legs, followed by up to four legless segments. As they grow, they continually moult, adding further segments and legs as they do so. Some species moult within specially prepared chambers of soil or silk, and may also shelter in these during wet weather, and most species eat the discarded exoskeleton after moulting.
The adult stage, when individuals become reproductively mature, is generally reached in the final moult stage, which varies between species and orders, although some species continue to moult after adulthood. Furthermore, some species alternate between reproductive and non-reproductive stages after maturity, a phenomenon known as periodomorphosis, in which the reproductive structures regress during non-reproductive stages. Millipedes may live from one to ten years, depending on species.
Millipedes occur on all continents except Antarctica, and occupy almost all terrestrial habitats. Typically forest floor dwellers, they live in leaf litter, dead wood, or soil, with a preference for humid conditions.
In temperate zones, millipedes are most abundant in moist deciduous forests, and may reach densities of over 1,000 individuals per square metre. Other habitats include coniferous forests, caves, and alpine ecosystems.
Deserticolous millipedes, species evolved to live in the desert, like Orthoporus ornatus, may show adaptations like a waxy epicuticle and the ability of water uptake from unsaturated air. Some species can survive freshwater floods and live submerged underwater for up to 11 months. A few species occur near the seashore and can survive in somewhat salty conditions.
Most millipedes are detritivores and feed on decomposing vegetation, feces, or organic matter mixed with soil. They often play important roles in the breakdown and decomposition of plant litter: estimates of consumption rates for individual species range from 1 to 11 percent of all leaf litter, depending on species and region, and collectively millipedes may consume nearly all the leaf litter in a region.
The leaf litter is fragmented in the millipede gut and excreted as pellets of leaf fragments, algae, fungi, and bacteria, which facilitates decomposition by the microorganisms. Where earthworm populations are low in tropical forests, millipedes play an important role in facilitating microbial decomposition of the leaf litter.
Some millipedes are herbivorous, feeding on living plants, and some species can become serious pests of crops. Millipedes in the order Polyxenida graze algae from bark, and Platydesmida feed on fungi.
A few species are omnivorous or in Callipodidan and Chordeumatida occasionally carnivorous feeding on insects, centipedes, earthworms, or snails.
Some species have piercing mouth parts that allow them to suck up plant juices.
Millipedes are preyed on by a wide range of animals, including various reptiles, amphibians, birds, mammals, and insects. Due to their lack of speed and their inability to bite or sting, millipedes' primary defence mechanism is to curl into a tight coil – protecting their delicate legs inside an armoured exoskeleton.
Many species also emit various foul-smelling liquid secretions through microscopic holes called ozopores (the openings of "odoriferous" or "repugnatorial glands"), along the sides of their bodies as a secondary defence.
Among the many irritant and toxic chemicals found in these secretions are alkaloids, benzoquinones, phenols, terpenoids, and hydrogen cyanide. Some of these substances are caustic and can burn the exoskeleton of ants and other insect predators, and the skin and eyes of larger predators.
Primates such as capuchin monkeys and lemurs have been observed intentionally irritating millipedes in order to rub the chemicals on themselves to repel mosquitoes. Some of these defensive compounds also show antifungal activity.
The bristly millipedes (order Polyxenida) lack both an armoured exoskeleton and odiferous glands, and instead are covered in numerous bristles that in at least one species, Polyxenus fasciculatus, detach and entangle ants.
Some millipedes form mutualistic relationships with organisms of other species, in which both species benefit from the interaction, or commensal relationships, in which only one species benefits while the other is unaffected.
Many millipede species have commensal relationships with mites of the orders Mesostigmata and Astigmata. Many of these mites are believed to be phoretic rather than parasitic, which means that they use the millipede host as a means of dispersal.
A novel interaction between millipedes and mosses was described in 2011, in which individuals of the newly discovered Psammodesmus bryophorus was found to have up to ten species living on its dorsal surface, in what may provide camouflage for the millipede and increased dispersal for the mosses.
Phylum: Arthropoda
Subphylum: Myriapoda
Class: Diplopoda
Blainville in Gervais, 1844
Genus: Helicorthomorpha
Species: H. holstii
Binomial name: Helicorthomorpha holstii
(Pocock, 1895)
Each double-legged segment is a result of two single segments fused together. Most millipedes have very elongated cylindrical or flattened bodies with more than 20 segments, while pill millipedes are shorter and can roll into a ball. Although the name "millipede" derives from the Latin for "thousand feet", no known species has 1,000; the record of 750 legs belongs to Illacme plenipes.
There are approximately 12,000 named species
classified into 16 orders and around 140 families, making Diplopoda the largest class of myriapods, an arthropod group which also includes centipedes and other multi-legged creatures.
Millipedes come in a variety of body shapes and sizes, ranging from 2 mm (0.08 in) to around 35 cm (14 in) in length and can have as few as eleven to over a hundred segments. They are generally black or brown in colour, although there are a few brightly coloured species, and some have aposematic colouring to warn that they are toxic, species of Motyxia produce cyanide as a chemical defence and are bioluminescent.
Body styles vary greatly between major millipede groups. In the basal subclass Penicillata, consisting of the tiny bristle millipedes, the exoskeleton is soft and uncalcified, and is covered in prominent setae or bristles.
All other millipedes, belonging to the subclass Chilognatha, have a hardened exoskeleton. The chilognaths are in turn divided into two infraclasses: the Pentazonia, containing relatively short-bodied groups such as pill millipedes, and the Helminthomorpha ("worm-like" millipedes), which contains the vast majority of species, with long, many-segmented bodies.
The head of a millipede is typically rounded above and flattened below and bears a pair of large mandibles in front of a plate-like structure called a gnathochilarium ("jaw lip").
The head contains a single pair of antennae with seven or eight segments and a group of sensory cones at the tip. Many orders also possess a pair of sensory organs known as the Tömösváry organs, shaped as small oval rings posterior and lateral to the base of the antennae. Their function is unknown, but they also occur in some centipedes, and are possibly used to measure humidity or light levels in the surrounding environment.
Millipede eyes consist of several simple flat lensed
nocelli arranged in a group or patch on each side of the head. These patches are also called ocular fields or ocellaria. Many species of millipedes, including the entire orders Polydesmida, Siphoniulida,
Glomeridesmida, Siphonophorida and Platydesmida, and cave-dwelling millipedes such as Causeyellan and Trichopetalum, had ancestors that could see but have subsequently lost their eyes and are blind.
Millipede bodies may be flattened or cylindrical, and are composed of numerous metameric segments, each with an exoskeleton consisting of four chitinous plates: a single plate above (the tergite), one at each side (pleurites), and a plate on the underside (sternite) where the legs attach.
In many millipedes, such as Merocheta and Juliformia, these plates are fused to varying degrees, sometimes forming a single cylindrical ring. The plates are typically hard, being impregnated with calcium salts. Because they can't close their permanently open spiracles and most species lack a waxy cuticle, millipedes are susceptible to water loss and with a few exceptions must spend most of their time in moist or humid environments.
The first segment behind the head is legless and known as a collum (from the Latin for neck or collar).
The second, third, and fourth body segments bear a single pair of legs each and are known as "haplosegments" (the three haplosegments are sometimes referred to as a "thorax").
The remaining segments, from the fifth to the posterior, are properly known as diplosegments or double segments, formed by the fusion of two embryonic segments. Each diplosegment bears two pairs of legs, rather than just one as in centipedes.
In some millipedes, the last few segments may be legless. The terms "segment" or "body ring" are often used interchangeably to refer to both haplo- and diplosegments. The final segment is known as the telson and consists of a legless preanal ring, a pair of anal valves (closeable plates around the anus), and a small scale below the anus.
Millipedes in several orders have keel-like extensions of the body-wall known as paranota, which can vary widely in shape, size, and texture; modifications include lobes, papillae, ridges, crests, spines and notches. Paranota may allow millipedes to wedge more securely into crevices, protect the legs, or make the millipede more difficult for predators to swallow.
The legs are composed of seven segments, and attach on the underside of the body. The legs of an individual are generally rather similar to each other, although often longer in males than females, and males of some species may have a reduced or enlarged first pair of legs.
The most conspicuous leg modifications are involved in reproduction, discussed below. Despite the common name, no millipede has been discovered with 1,000 legs: common species have between 34 and 400 legs, and the record is held by Illacme plenipes, with individuals possessing up to 750 legs – more than any other creature on Earth.
Millipedes breathe through two pairs of spiracles located ventrally on each segment near the base of the legs. Each opens into an internal pouch, and connects to a system of tracheae. The heart runs the entire length of the body, with an aorta stretching into the head. The excretory organs are two pairs of malpighian tubules, located near the mid-part of the gut. The digestive tract is a simple tube with two pairs of salivary glands to help digest the food.
Millipedes show a diversity of mating styles and structures. In the basal order Polyxenida (bristle millipedes), mating is indirect: males deposit spermatophores onto webs they secrete with special glands, and the spermatophores are subsequently picked up by females.
In all other millipede groups, males possess one or two pairs of modified legs called gonopods which are used to transfer sperm to the female during copulation.
The location of the gonopods differs between groups: in males of the Pentazonia they are located at the rear of the body and known as telopods and may also function in grasping females, while in the Helminthomorpha – the vast majority of species – they are located on the seventh body segment. A few species are parthenogenetic, having few, if any, males.
Gonopods occur in a diversity of shapes and sizes, and in the range from closely resembling walking legs to complex structures quite unlike legs at all.
In some groups, the gonopods are kept retracted within the body; in others they project forward parallel to the body. Gonopod morphology is the predominant means of determining species among millipedes: the structures may differ greatly between closely related species but very little within a species. The gonopods develop gradually from walking legs through successive moults until reproductive maturity.
The genital openings (gonopores) of both sexes are located on the underside of the third body segment (near the second pair of legs) and may be accompanied in the male by one or two penes which deposit the sperm packets onto the gonopods.
In the female, the genital pores open into paired small sacs called cyphopods or vulvae, which are covered by small hood-like lids, and are used to store the sperm after copulation. The cyphopod morphology can also be used to identify species. Millipede sperm lack flagella, a unique trait among myriapods.
In all except the bristle millipedes, copulation occurs with the two individuals facing one another.
Copulation may be preceded by male behaviours such as tapping with antennae, running along the back of the female, offering edible glandular secretions, or in the case of some pill-millipedes, stridulation or "chirping". During copulation in most millipedes, the male positions his seventh segment in front of the female's third segment, and may insert his gonopods to extrude the vulvae before bending his body to deposit sperm onto his gonopods and reinserting the "charged" gonopods into the female.
Females lay from ten to three hundred eggs at a time, depending on species, fertilising them with the stored sperm as they do so. Many species deposit the eggs on moist soil or organic detritus, but some construct nests lined with dried faeces, and may protect the eggs within silk cocoons. In most species, the female abandons the eggs after they are laid, but some species in the orders Platydesmida and Stemmiulida provide parental care for eggs and young.
The young hatch after a few weeks, and typically have only three pairs of legs, followed by up to four legless segments. As they grow, they continually moult, adding further segments and legs as they do so. Some species moult within specially prepared chambers of soil or silk, and may also shelter in these during wet weather, and most species eat the discarded exoskeleton after moulting.
The adult stage, when individuals become reproductively mature, is generally reached in the final moult stage, which varies between species and orders, although some species continue to moult after adulthood. Furthermore, some species alternate between reproductive and non-reproductive stages after maturity, a phenomenon known as periodomorphosis, in which the reproductive structures regress during non-reproductive stages. Millipedes may live from one to ten years, depending on species.
Millipedes occur on all continents except Antarctica, and occupy almost all terrestrial habitats. Typically forest floor dwellers, they live in leaf litter, dead wood, or soil, with a preference for humid conditions.
In temperate zones, millipedes are most abundant in moist deciduous forests, and may reach densities of over 1,000 individuals per square metre. Other habitats include coniferous forests, caves, and alpine ecosystems.
Deserticolous millipedes, species evolved to live in the desert, like Orthoporus ornatus, may show adaptations like a waxy epicuticle and the ability of water uptake from unsaturated air. Some species can survive freshwater floods and live submerged underwater for up to 11 months. A few species occur near the seashore and can survive in somewhat salty conditions.
Most millipedes are detritivores and feed on decomposing vegetation, feces, or organic matter mixed with soil. They often play important roles in the breakdown and decomposition of plant litter: estimates of consumption rates for individual species range from 1 to 11 percent of all leaf litter, depending on species and region, and collectively millipedes may consume nearly all the leaf litter in a region.
The leaf litter is fragmented in the millipede gut and excreted as pellets of leaf fragments, algae, fungi, and bacteria, which facilitates decomposition by the microorganisms. Where earthworm populations are low in tropical forests, millipedes play an important role in facilitating microbial decomposition of the leaf litter.
Some millipedes are herbivorous, feeding on living plants, and some species can become serious pests of crops. Millipedes in the order Polyxenida graze algae from bark, and Platydesmida feed on fungi.
A few species are omnivorous or in Callipodidan and Chordeumatida occasionally carnivorous feeding on insects, centipedes, earthworms, or snails.
Some species have piercing mouth parts that allow them to suck up plant juices.
Millipedes are preyed on by a wide range of animals, including various reptiles, amphibians, birds, mammals, and insects. Due to their lack of speed and their inability to bite or sting, millipedes' primary defence mechanism is to curl into a tight coil – protecting their delicate legs inside an armoured exoskeleton.
Many species also emit various foul-smelling liquid secretions through microscopic holes called ozopores (the openings of "odoriferous" or "repugnatorial glands"), along the sides of their bodies as a secondary defence.
Among the many irritant and toxic chemicals found in these secretions are alkaloids, benzoquinones, phenols, terpenoids, and hydrogen cyanide. Some of these substances are caustic and can burn the exoskeleton of ants and other insect predators, and the skin and eyes of larger predators.
Primates such as capuchin monkeys and lemurs have been observed intentionally irritating millipedes in order to rub the chemicals on themselves to repel mosquitoes. Some of these defensive compounds also show antifungal activity.
The bristly millipedes (order Polyxenida) lack both an armoured exoskeleton and odiferous glands, and instead are covered in numerous bristles that in at least one species, Polyxenus fasciculatus, detach and entangle ants.
Some millipedes form mutualistic relationships with organisms of other species, in which both species benefit from the interaction, or commensal relationships, in which only one species benefits while the other is unaffected.
Many millipede species have commensal relationships with mites of the orders Mesostigmata and Astigmata. Many of these mites are believed to be phoretic rather than parasitic, which means that they use the millipede host as a means of dispersal.
A novel interaction between millipedes and mosses was described in 2011, in which individuals of the newly discovered Psammodesmus bryophorus was found to have up to ten species living on its dorsal surface, in what may provide camouflage for the millipede and increased dispersal for the mosses.
Phylum: Arthropoda
Subphylum: Myriapoda
Class: Diplopoda
Blainville in Gervais, 1844
Genus: Helicorthomorpha
Species: H. holstii
Binomial name: Helicorthomorpha holstii
(Pocock, 1895)


Opisthodolichopus scandens

Millipedes are among the first animals to have colonised land during the Silurian period. Early forms probably ate mosses and primitive vascular plants.
There are two major groups of millipedes whose members are all extinct: the Archipolypoda ("ancient, many-legged ones") which contain the oldest known terrestrial animals, and Arthropleuridea, which contain the largest known land invertebrates.
Millipedes are a group of arthropods that are characterised by having two pairs of jointed legs on most body segments; they are known scientifically as the class Diplopoda, the name being derived from this feature. Each double-legged segment is a result of two single segments fused together while centipedes have one pair per segment.
The differences between millipedes and centipedes are common question from the general public.
Both group of myriapods share similarities in long multi-segmented bodies, many legs, a single pair of antennae and the presence of posyanntennal organs.
The head alone exemplifies the differences, millipedes have short geniculate (elbowed) antennae for probing the substrate, a pair of robust mandibles and a single pair of maxillae fused into a lip, while centipedes have long threadlike antennae, a pair of small mandibles, two pairs of maxillae and a pair of large poison ckaws.
Also most millipedes have very elongated cylindrical or flattened bodies with more than 20 segments, while pill millipedes are shorter and can roll into a ball.
The name "millipede" derives from the Latin for "thousand feet", no known species has 1,000; the record of 750 legs belongs to Illacme plenipes.
There are approximately 12,000 named species classified into 16 orders and around 140 families, making Diplopoda the largest class of myriapods, an arthropod group which also includes centipedes and other multi-legged creatures.
Most millipedes are slow-moving detritivores, eating decaying leaves and other dead plant matter. Some eat fungi or suck plant fluids, and a small minority are predatory. Millipedes are generally harmless to humans, although some can become household or garden pests.
Millipedes can be unwanted especially in greenhouses where they can cause severe damage to emergent seedlings. Most millipedes defend themselves with a variety of chemicals secreted from pores along the body, although the tiny bristle millipedes are covered with tufts of detachable bristles. Its primary defence mechanism is to curl into a tight coil, thereby protecting its legs and other vital delicate areas on the body behind a hard exoskeleton. Reproduction in most species is carried out by modified male legs called gonopods, which transfer packets of sperm to females.
Among myriapods, millipedes have traditionally been considered most closely related to the tiny pauropods, although some molecular studies challenge this relationship. Millipedes can be distinguished from the somewhat similar but only distantly related centipedes (class Chilopoda), which move rapidly, are venomous, carnivorous, and have only a single pair of legs on each body segment.
The scientific study of millipedes is known as diplopodology, and a scientist who studies them is called a diplopodologist.
Approximately 12,000 millipede species have been described. Estimates of the true number of species on earth range from 15,000 to as high as 80,000. Few species of millipede are at all widespread; they have very poor dispersal abilities, depending as they do on terrestrial locomotion and humid habitats. These factors have favoured genetic isolation and rapid speciation, producing many lineages with restricted ranges.
The living members of the Diplopoda are divided into sixteen orders in two subclasses. The basal subclass Penicillata contains a single order, Polyxenida (bristle millipedes).
All other millipedes belong to the subclass Chilognatha consisting of two infraclasses: Pentazonia, containing the short-bodied pill millipedes, and Helminthomorpha (worm-like millipedes), containing the great majority of the species.
Opisthodolichopus scandens are easily recognised by their long segmented body with a dark line on the back and with two pairs of legs on most segments, except for the first few and last segments . The name means “a thousand legs”, but while there are species in the world with more than 700 legs, the ones seen in Singapore seldom have more than 100.
As they grow, they need to moult discard the old skeleton and grow a new one. Their cuticle is not waterproof, and hence they are largely terrestrial.
Most species are also nocturnal to avoid water loss and are most readily found in damp habitats such as the leaf litter or underground and they feed on decaying organic matter or fungi, and hence they are commonly seen in the leaf litter or on fallen trees. They can breathe by taking in air through openings in the cuticle.
Most species are not venomous, and protect themselves either by secreting toxic or distasteful chemicals, or roll up to protect their softer underparts, exposing only the tougher upperparts.
To breed, the males usually twist their bodies around the females to transfer the sperm, and the females will lay the eggs inside a nest in the soil. The newly hatched millipedes usually have much fewer legs than the adults, and gain more legs and body segments as they moult.
Phylum: Arthropoda
Subphylum: Myriapoda
Class: Diplopoda
Blainville in Gervais, 1844
Genus: Opisthodolichopus
Species: O.scandens
Binomial name: Opisthodolichopus scandens
Hoffman 1973
There are two major groups of millipedes whose members are all extinct: the Archipolypoda ("ancient, many-legged ones") which contain the oldest known terrestrial animals, and Arthropleuridea, which contain the largest known land invertebrates.
Millipedes are a group of arthropods that are characterised by having two pairs of jointed legs on most body segments; they are known scientifically as the class Diplopoda, the name being derived from this feature. Each double-legged segment is a result of two single segments fused together while centipedes have one pair per segment.
The differences between millipedes and centipedes are common question from the general public.
Both group of myriapods share similarities in long multi-segmented bodies, many legs, a single pair of antennae and the presence of posyanntennal organs.
The head alone exemplifies the differences, millipedes have short geniculate (elbowed) antennae for probing the substrate, a pair of robust mandibles and a single pair of maxillae fused into a lip, while centipedes have long threadlike antennae, a pair of small mandibles, two pairs of maxillae and a pair of large poison ckaws.
Also most millipedes have very elongated cylindrical or flattened bodies with more than 20 segments, while pill millipedes are shorter and can roll into a ball.
The name "millipede" derives from the Latin for "thousand feet", no known species has 1,000; the record of 750 legs belongs to Illacme plenipes.
There are approximately 12,000 named species classified into 16 orders and around 140 families, making Diplopoda the largest class of myriapods, an arthropod group which also includes centipedes and other multi-legged creatures.
Most millipedes are slow-moving detritivores, eating decaying leaves and other dead plant matter. Some eat fungi or suck plant fluids, and a small minority are predatory. Millipedes are generally harmless to humans, although some can become household or garden pests.
Millipedes can be unwanted especially in greenhouses where they can cause severe damage to emergent seedlings. Most millipedes defend themselves with a variety of chemicals secreted from pores along the body, although the tiny bristle millipedes are covered with tufts of detachable bristles. Its primary defence mechanism is to curl into a tight coil, thereby protecting its legs and other vital delicate areas on the body behind a hard exoskeleton. Reproduction in most species is carried out by modified male legs called gonopods, which transfer packets of sperm to females.
Among myriapods, millipedes have traditionally been considered most closely related to the tiny pauropods, although some molecular studies challenge this relationship. Millipedes can be distinguished from the somewhat similar but only distantly related centipedes (class Chilopoda), which move rapidly, are venomous, carnivorous, and have only a single pair of legs on each body segment.
The scientific study of millipedes is known as diplopodology, and a scientist who studies them is called a diplopodologist.
Approximately 12,000 millipede species have been described. Estimates of the true number of species on earth range from 15,000 to as high as 80,000. Few species of millipede are at all widespread; they have very poor dispersal abilities, depending as they do on terrestrial locomotion and humid habitats. These factors have favoured genetic isolation and rapid speciation, producing many lineages with restricted ranges.
The living members of the Diplopoda are divided into sixteen orders in two subclasses. The basal subclass Penicillata contains a single order, Polyxenida (bristle millipedes).
All other millipedes belong to the subclass Chilognatha consisting of two infraclasses: Pentazonia, containing the short-bodied pill millipedes, and Helminthomorpha (worm-like millipedes), containing the great majority of the species.
Opisthodolichopus scandens are easily recognised by their long segmented body with a dark line on the back and with two pairs of legs on most segments, except for the first few and last segments . The name means “a thousand legs”, but while there are species in the world with more than 700 legs, the ones seen in Singapore seldom have more than 100.
As they grow, they need to moult discard the old skeleton and grow a new one. Their cuticle is not waterproof, and hence they are largely terrestrial.
Most species are also nocturnal to avoid water loss and are most readily found in damp habitats such as the leaf litter or underground and they feed on decaying organic matter or fungi, and hence they are commonly seen in the leaf litter or on fallen trees. They can breathe by taking in air through openings in the cuticle.
Most species are not venomous, and protect themselves either by secreting toxic or distasteful chemicals, or roll up to protect their softer underparts, exposing only the tougher upperparts.
To breed, the males usually twist their bodies around the females to transfer the sperm, and the females will lay the eggs inside a nest in the soil. The newly hatched millipedes usually have much fewer legs than the adults, and gain more legs and body segments as they moult.
Phylum: Arthropoda
Subphylum: Myriapoda
Class: Diplopoda
Blainville in Gervais, 1844
Genus: Opisthodolichopus
Species: O.scandens
Binomial name: Opisthodolichopus scandens
Hoffman 1973



Nemertea
Terrestrial ribbon worm
Terrestrial ribbon worm

Nemertea is a phylum of invertebrat animals also
known as ribbon worms or proboscis worms.
Alternative names for the phylum have included Nemertini, Nemertinea and Rhynchocoela.
Most are very slim, usually only a few millimeters wide, although a few have relatively short but wide bodies. Many have patterns of yellow, orange, red and green coloration. The foregut, stomach and intestine run a little below the midline of the body, the anus is at the tip of the tail, and the mouth is under the front. A little above the gut is the rhynchocoel, a cavity which mostly runs above the midline and ends a little short of the rear of the body.
All species have a proboscis which lies in the rhynchocoel when inactive but everts (turns inside-out) to emerge just above the mouth and capture the animal's prey with venom. A highly extensible muscle in the back of the rhynchocoel pulls the proboscis in when an attack ends. A few species with stubby bodies filter feed and have suckers at the front and back ends, with which they attach to a host.
The brain is a ring of four ganglia, positioned around the rhynchocoel near its front end. At least a pair of ventral nerve cords connect to the brain and run along the length of the body.
Most nemerteans have various chemoreceptors, and on their heads some species have a number of pigment-cup ocelli. These ocelli can detect light but can not form an image.
Nemerteans respire through the skin. They have at least two lateral vessels which are joined at the ends to form a loop, and these and the rhynchocoel are filled with fluid. There is no heart, and the flow of fluid depends on contraction of muscles in the vessels and the body wall. To filter out soluble waste products, flame cells are embedded in the front part of the two lateral fluid vessels, and remove the wastes through a network of pipes to the outside.
All nemerteans move slowly, using their external cilia to glide on surfaces on a trail of slime, while larger species use muscular waves to crawl, and some swim by dorso-ventral undulations.
A few live in the open ocean while the rest find or make hiding places on the bottom. About a dozen species inhabit freshwater, mainly in the tropics and subtropics, and another dozen species live on land in cool, damp places. Most nemerteans are carnivores, feeding on annelids, clams and crustaceans. A few species are scavengers, and a few species live commensally inside the mantle cavity of molluscs. Some species have devastated commercial fishing of clams and crabs. Nemerteans have few predators. Two species are sold as fish bait.
In most species the sexes are separate, but all the freshwater species are hermaphroditic. Nemerteans often have numerous temporary gonads (ovaries or testes), and build temporary gonoducts (ducts from which the ova or sperm are emitted) opening to a gonopore, one per gonad, when the ova and sperm are ready.
The eggs are generally fertilised externally. Some species shed them into the water, and others protect their eggs in various ways. The fertilized egg divides by spiral cleavage and grows by determinate development, in which the fate of a cell can usually be predicted from its predecessors in the process of division. The embryos of most taxa develop either directly to form juveniles (like the adult but smaller) or to form planuliform larvae, in which the larva's long axis is the same as the juvenile's. However, some form a pilidium larva, in which the developing juvenile has a gut which lies across the larva's body, and usually eats the remains of the larva when it emerges. The bodies of some species fragment readily, and even parts near the tail can grow full bodies.
It has been suggested that three fossil species may be nemerteans, but none is confirmed. Traditional taxonomy divides the phylum in two classes, Anopla ("unarmed" – their proboscises do not have a little dagger) with two orders, and Enopla ("armed" with a dagger) with two orders. However, it is now accepted that Anopla are paraphyletic (have given rise to another group), as one order is more closely related to Enopla than to the other order of Anopla.
The phylum Nemertea is monophyletic. Its synapomorphies include the rhynchocoel and eversible proboscis. Traditional taxonomy says that nemerteans are closely related to flatworms. Both phyla are regarded as members of the
Lophotrochozoa, a very large "super-phylum" that also includes molluscs, annelids, brachiopods, bryozoa and many other protostomes.
Nemerteans are very unusual animals. The typical nemertean body is very slim in proportion to its length. The smallest are a few millimeters long, most are less than 20 centimetres (7.9 in), and several exceed 1 metre (3.3 ft). The longest animal ever found, at 54 metres (177 ft) long, may be a specimen of Lineus longissimus.
[Note] although L. longissimus is usually only a few millimeters wide. The bodies of most nemerteans can stretch a lot, up to 10 times their resting length in some species, but reduce their length to 50% and increase their width to 300% when disturbed. A few have relatively short but wide bodies,
Smaller nemerteans are approximately cylindrical, but larger species are flattened dorso-ventrally. Many have visible patterns in various combinations of yellow, orange, red and green.
The outermost layer of the body has no cuticle but consists of a ciliated and glandular epithelium containing rhabdites, which form the mucus in which the cilia glide. Each ciliated cell has many cilia and microvilli. The outermost layer rests on a thickened basement membrane, the dermis. Next to the dermis are at least three layers of muscles, some circular and some longitudinal. The combinations of muscle types vary between the different classes, but these are not associated with differences in movement. Nemerteans also have dorso-ventral muscles, which flatten the animals, especially in the larger species. Inside the concentric tubes of these layers is mesenchyme, a kind of connective tissue. In pelagic species this tissue is gelatinous and buoyant.
Kingdom: Animalia
Subkingdom: Eumetazoa
Clade: ParaHoxozoa
Clade: Bilateria
Clade: Nephrozoa
(unranked): Protostomia
(unranked): Spiralia
Superphylum: Lophotrochozoa
Phylum: Nemertea
Schultze, 1851
known as ribbon worms or proboscis worms.
Alternative names for the phylum have included Nemertini, Nemertinea and Rhynchocoela.
Most are very slim, usually only a few millimeters wide, although a few have relatively short but wide bodies. Many have patterns of yellow, orange, red and green coloration. The foregut, stomach and intestine run a little below the midline of the body, the anus is at the tip of the tail, and the mouth is under the front. A little above the gut is the rhynchocoel, a cavity which mostly runs above the midline and ends a little short of the rear of the body.
All species have a proboscis which lies in the rhynchocoel when inactive but everts (turns inside-out) to emerge just above the mouth and capture the animal's prey with venom. A highly extensible muscle in the back of the rhynchocoel pulls the proboscis in when an attack ends. A few species with stubby bodies filter feed and have suckers at the front and back ends, with which they attach to a host.
The brain is a ring of four ganglia, positioned around the rhynchocoel near its front end. At least a pair of ventral nerve cords connect to the brain and run along the length of the body.
Most nemerteans have various chemoreceptors, and on their heads some species have a number of pigment-cup ocelli. These ocelli can detect light but can not form an image.
Nemerteans respire through the skin. They have at least two lateral vessels which are joined at the ends to form a loop, and these and the rhynchocoel are filled with fluid. There is no heart, and the flow of fluid depends on contraction of muscles in the vessels and the body wall. To filter out soluble waste products, flame cells are embedded in the front part of the two lateral fluid vessels, and remove the wastes through a network of pipes to the outside.
All nemerteans move slowly, using their external cilia to glide on surfaces on a trail of slime, while larger species use muscular waves to crawl, and some swim by dorso-ventral undulations.
A few live in the open ocean while the rest find or make hiding places on the bottom. About a dozen species inhabit freshwater, mainly in the tropics and subtropics, and another dozen species live on land in cool, damp places. Most nemerteans are carnivores, feeding on annelids, clams and crustaceans. A few species are scavengers, and a few species live commensally inside the mantle cavity of molluscs. Some species have devastated commercial fishing of clams and crabs. Nemerteans have few predators. Two species are sold as fish bait.
In most species the sexes are separate, but all the freshwater species are hermaphroditic. Nemerteans often have numerous temporary gonads (ovaries or testes), and build temporary gonoducts (ducts from which the ova or sperm are emitted) opening to a gonopore, one per gonad, when the ova and sperm are ready.
The eggs are generally fertilised externally. Some species shed them into the water, and others protect their eggs in various ways. The fertilized egg divides by spiral cleavage and grows by determinate development, in which the fate of a cell can usually be predicted from its predecessors in the process of division. The embryos of most taxa develop either directly to form juveniles (like the adult but smaller) or to form planuliform larvae, in which the larva's long axis is the same as the juvenile's. However, some form a pilidium larva, in which the developing juvenile has a gut which lies across the larva's body, and usually eats the remains of the larva when it emerges. The bodies of some species fragment readily, and even parts near the tail can grow full bodies.
It has been suggested that three fossil species may be nemerteans, but none is confirmed. Traditional taxonomy divides the phylum in two classes, Anopla ("unarmed" – their proboscises do not have a little dagger) with two orders, and Enopla ("armed" with a dagger) with two orders. However, it is now accepted that Anopla are paraphyletic (have given rise to another group), as one order is more closely related to Enopla than to the other order of Anopla.
The phylum Nemertea is monophyletic. Its synapomorphies include the rhynchocoel and eversible proboscis. Traditional taxonomy says that nemerteans are closely related to flatworms. Both phyla are regarded as members of the
Lophotrochozoa, a very large "super-phylum" that also includes molluscs, annelids, brachiopods, bryozoa and many other protostomes.
Nemerteans are very unusual animals. The typical nemertean body is very slim in proportion to its length. The smallest are a few millimeters long, most are less than 20 centimetres (7.9 in), and several exceed 1 metre (3.3 ft). The longest animal ever found, at 54 metres (177 ft) long, may be a specimen of Lineus longissimus.
[Note] although L. longissimus is usually only a few millimeters wide. The bodies of most nemerteans can stretch a lot, up to 10 times their resting length in some species, but reduce their length to 50% and increase their width to 300% when disturbed. A few have relatively short but wide bodies,
Smaller nemerteans are approximately cylindrical, but larger species are flattened dorso-ventrally. Many have visible patterns in various combinations of yellow, orange, red and green.
The outermost layer of the body has no cuticle but consists of a ciliated and glandular epithelium containing rhabdites, which form the mucus in which the cilia glide. Each ciliated cell has many cilia and microvilli. The outermost layer rests on a thickened basement membrane, the dermis. Next to the dermis are at least three layers of muscles, some circular and some longitudinal. The combinations of muscle types vary between the different classes, but these are not associated with differences in movement. Nemerteans also have dorso-ventral muscles, which flatten the animals, especially in the larger species. Inside the concentric tubes of these layers is mesenchyme, a kind of connective tissue. In pelagic species this tissue is gelatinous and buoyant.
Kingdom: Animalia
Subkingdom: Eumetazoa
Clade: ParaHoxozoa
Clade: Bilateria
Clade: Nephrozoa
(unranked): Protostomia
(unranked): Spiralia
Superphylum: Lophotrochozoa
Phylum: Nemertea
Schultze, 1851

Scolopendra subspinipes


Scolopendra subspinipes is a species of very large centipede found throughout eastern Asia.
One of the most widespread and common species in the genus Scolopendra, this species is also found on virtually all land areas around and within the Indian Ocean, all of tropical and subtropical Asia from Russia to the islands of Malaysia and Indonesia, Singapore, Australia, South and Central America, the Caribbean islands, and possibly parts of the southern United States.
However, how much of this range is natural and how much due to human introduction is unclear. With a wide geographic range and numerous color variations, the species is known by a great many common names including Chinese red-headed centipede, jungle centipede, orange-legged centipede, red-headed centipede and Vietnamese centipede.
It is among the largest centipedes with a maximum length of 20 cm. This centipede is an active, aggressive predator that preys on any animal it can overpower.
This is a large species which can grow up to 20 cm in length. It has colour variations. Its body is usually red or reddish brown with yellow or yellow-orange legs. In common with other members of genus Scolopendra, it has 22 body segments with each segment having one pair of legs.
A pair of modified legs known as forcipules can be found on its head, which is covered by a flat shield and bears a pair of antennae. The forcipules are the major tools used by the centipede to kill its prey or for defense, as they have sharp claws that connect to venom glands.
Centipedes breathe through spiracles (the openings located along the sides of their bodies). These openings are either round-shaped or S-shaped. They have simple eyes with poor vision, so they rely much on touch and their chemoreceptors.
The species can be found in tropical and subtropical regions throughout the Old World.
This is an aggressive and nervous arthropod which is ready to strike if interfered with and is sensitive to vibrations nearby.
It preys primarily on arachnids, including spiders, scorpions, and vinegaroons. However, if it is large enough to overpower small vertebrates like mice or small reptiles, it will readily attempt to consume them as well. It tends to try to eat almost every living animal it encounters that is not longer than itself. It seizes prey with its anterior legs, then curves its head quickly behind to implant its venomous jaws deeply and firmly into the prey. The prey is held by the centipede's other legs until it dies from the fast-acting venom. During a fight, the centipede will use its entire body coiling the prey or enemy with its legs firmly attaching to the body of the opponent. Then, it will quickly penetrate its forcipules into the victim for venom injection.
The male produces capsules containing mature sperm cells, spermatophores, which are deposited in a reservoir called the spermathecae of the female during mating.
The female then fertilizes her immature eggs, oocytes, and deposits them in a dark, protected area. The female lays 50 to 80 eggs which she vigilantly protects until they hatch and the baby centipede molt once. If danger is detected she will wrap around her babies to keep them safe. The young centipedes molt once each year, and take three to four years to attain full adult size. Adults molt once every year. They may live for 10 years or more.
Scolopendra subspinipes has been reported as the apparent cause of a human death. The fatal case was in Philippines in which the centipede bit a seven-year-old girl on her head. She died 29 hours later.
S. subspinipes is a popular pet among arthropod hobbyists. The centipede was a traditional food source for Aboriginal Australians.
The number of subspecies of S. subspinipes is unclear and varies between authors.
Taxonomic characters have incorporated plastic traits such as color and sulcus structure and the number and position of spines, producing indistinguishable and intergrading subspecies.
A 2012 review found that one former subspecies, S. subspinipes cingulatoides is in fact a distinct species, and that S. subspinipes has no valid subspecies.
Phylum: Arthropoda
Class: Chilopoda
Order: Scolopendromorpha
Family: Scolopendre
Genus: Scolopendra
Species: S. subspinipes
Binomial nameb Scolopendra subspinipes
Leach, 1815
One of the most widespread and common species in the genus Scolopendra, this species is also found on virtually all land areas around and within the Indian Ocean, all of tropical and subtropical Asia from Russia to the islands of Malaysia and Indonesia, Singapore, Australia, South and Central America, the Caribbean islands, and possibly parts of the southern United States.
However, how much of this range is natural and how much due to human introduction is unclear. With a wide geographic range and numerous color variations, the species is known by a great many common names including Chinese red-headed centipede, jungle centipede, orange-legged centipede, red-headed centipede and Vietnamese centipede.
It is among the largest centipedes with a maximum length of 20 cm. This centipede is an active, aggressive predator that preys on any animal it can overpower.
This is a large species which can grow up to 20 cm in length. It has colour variations. Its body is usually red or reddish brown with yellow or yellow-orange legs. In common with other members of genus Scolopendra, it has 22 body segments with each segment having one pair of legs.
A pair of modified legs known as forcipules can be found on its head, which is covered by a flat shield and bears a pair of antennae. The forcipules are the major tools used by the centipede to kill its prey or for defense, as they have sharp claws that connect to venom glands.
Centipedes breathe through spiracles (the openings located along the sides of their bodies). These openings are either round-shaped or S-shaped. They have simple eyes with poor vision, so they rely much on touch and their chemoreceptors.
The species can be found in tropical and subtropical regions throughout the Old World.
This is an aggressive and nervous arthropod which is ready to strike if interfered with and is sensitive to vibrations nearby.
It preys primarily on arachnids, including spiders, scorpions, and vinegaroons. However, if it is large enough to overpower small vertebrates like mice or small reptiles, it will readily attempt to consume them as well. It tends to try to eat almost every living animal it encounters that is not longer than itself. It seizes prey with its anterior legs, then curves its head quickly behind to implant its venomous jaws deeply and firmly into the prey. The prey is held by the centipede's other legs until it dies from the fast-acting venom. During a fight, the centipede will use its entire body coiling the prey or enemy with its legs firmly attaching to the body of the opponent. Then, it will quickly penetrate its forcipules into the victim for venom injection.
The male produces capsules containing mature sperm cells, spermatophores, which are deposited in a reservoir called the spermathecae of the female during mating.
The female then fertilizes her immature eggs, oocytes, and deposits them in a dark, protected area. The female lays 50 to 80 eggs which she vigilantly protects until they hatch and the baby centipede molt once. If danger is detected she will wrap around her babies to keep them safe. The young centipedes molt once each year, and take three to four years to attain full adult size. Adults molt once every year. They may live for 10 years or more.
Scolopendra subspinipes has been reported as the apparent cause of a human death. The fatal case was in Philippines in which the centipede bit a seven-year-old girl on her head. She died 29 hours later.
S. subspinipes is a popular pet among arthropod hobbyists. The centipede was a traditional food source for Aboriginal Australians.
The number of subspecies of S. subspinipes is unclear and varies between authors.
Taxonomic characters have incorporated plastic traits such as color and sulcus structure and the number and position of spines, producing indistinguishable and intergrading subspecies.
A 2012 review found that one former subspecies, S. subspinipes cingulatoides is in fact a distinct species, and that S. subspinipes has no valid subspecies.
Phylum: Arthropoda
Class: Chilopoda
Order: Scolopendromorpha
Family: Scolopendre
Genus: Scolopendra
Species: S. subspinipes
Binomial nameb Scolopendra subspinipes
Leach, 1815




Platerodrilus ruficollis
Trilobite beetle
Trilobite beetle

The trilobite beetle, a creature that looks like it’d be more at home with the dinosaurs than in the forests of present day.
Found in Southeast Asia and India, these peculiar beetles aren’t related to trilobites, but they are named after them, because of their similarly armoured bodies and helmet-like heads.
And their strange looks are just the beginning. Their mating habits are so mysterious, they’ve kept scientists guessing for more than two centuries
First discovered back in the 1800s, trilobite beetles belong to the family Lycidae, commonly referred to as net-winged beetles. A major characteristic of many net-winged beetle species is that their males and females look entirely different.
The most confounding thing about trilobite beetles – every single individual that looks like a trilobite is a female. That form is actually retained from when they were larvae, so the females change very little from when they hatch out of their eggs.
The males are different they basically look like plain old beetles, with long, winged bodies and a pair of thick antennae. And they tend to only grow to about 5 mm long, whereas the females can stretch to more than 6 cm.
Male trilobite beetles look like every other beetle you might see hiding out in the forest, the only way to confirm what one actually looks like is by catching it in the act of mating with a female.
Platerodrilus ruficollis is a species of trilobite beetle found in Malaysia and Singapore, also known by its junior synonym, Duliticola hoiseni.
Platerodrilus is a genus of beetles of the family Lycidae. They commonly appear in the literature under the name Duliticola, which is an obsolete junior synonym.
The females retain a larval form as adults and are about 40–80 mm in length. The females and larvae have a flattened, dark body with large scales over the head, resembling trilobites, hence the informal names trilobite beetle, trilobite larva or Sumatran trilobite larva.
In contrast, the males are much smaller, 8–9 mm, resembling other beetles. Species are found in tropical rainforests of India and South-east Asia.
Trilobites ( meaning "three lobes") are a group of extinct marine artiopodan arthropods that form the
class Trilobita.
Trilobites form one of the earliest-known groups of arthropods. The first appearance of trilobites in the fossil record defines the base of the Atdabanian stage of the Early Cambrian period (521 million years ago), and they flourished throughout the lower Paleozoic before slipping into a long decline, when, during the Devonian, all trilobite orders except the Proetida died out. The last extant trilobites finally disappeared in the mass extinction at the end of the Permian about 252 million years ago. Trilobites were among the most successful of all early animals, existing in oceans for almost 300 million years.
By the time trilobites first appeared in the fossil record, they were already highly diversified and geographically dispersed. Because trilobites had wide diversity and an easily fossilized exoskeleton, they left an extensive fossil record. The study of their fossils has facilitated important contributions to biostratigraphy, paleontology, evolutionary biology, and plate tectonics.
Trilobites are often placed within the arthropod subphylum Schizoramia within the superclass Arachnomorpha (equivalent to the Arachnata), although several alternative taxonomies are found in the literature. More recently they have been placed within the Artiopoda, which includes many organisms that are morphologically similar to trilobites, but are largely unmineralised.
Trilobites had many lifestyles; some moved over the seabed as predators, scavengers, or filter feeders, and some swam, feeding on plankton. Some even crawled onto land.
Most lifestyles expected of modern marine arthropods are seen in trilobites, with the possible exception of parasitism (where scientific debate continues). Some trilobites (particularly the family Olenidae) are even thought to have evolved a symbiotic relationship with sulfur-eating bacteria from which they derived food.
The largest trilobites were more than 45 centimetres (18 in) long and may have weighed as much as 4.5 kilograms (9.9 lb).
Surprisingly enough, even the world's leading experts on these beetles are not 100% sure what they feed on, and up until relatively recently, there was some conflict as to what their diet consisted of.
However, the most popular and widely accepted theory is that these beetles feed on microorganisms living within the juices of rotten logs, as outlined in the findings of Alvin T. C. Wong (1996). Most Platerodrilus experts agree with Wong on that, so for now it should be treated as fact. Besides theres information that this sp do feed on slime mold too.
Kingdom: Animalia
Phylum: Arthropoda
Class: Insecta
Order: Coleoptera
Family: Lycidae
Genus: Platerodrilus
Species: P. ruficollis
Binomial name Platerodrilus ruficollis
(Pic, 1942)
Found in Southeast Asia and India, these peculiar beetles aren’t related to trilobites, but they are named after them, because of their similarly armoured bodies and helmet-like heads.
And their strange looks are just the beginning. Their mating habits are so mysterious, they’ve kept scientists guessing for more than two centuries
First discovered back in the 1800s, trilobite beetles belong to the family Lycidae, commonly referred to as net-winged beetles. A major characteristic of many net-winged beetle species is that their males and females look entirely different.
The most confounding thing about trilobite beetles – every single individual that looks like a trilobite is a female. That form is actually retained from when they were larvae, so the females change very little from when they hatch out of their eggs.
The males are different they basically look like plain old beetles, with long, winged bodies and a pair of thick antennae. And they tend to only grow to about 5 mm long, whereas the females can stretch to more than 6 cm.
Male trilobite beetles look like every other beetle you might see hiding out in the forest, the only way to confirm what one actually looks like is by catching it in the act of mating with a female.
Platerodrilus ruficollis is a species of trilobite beetle found in Malaysia and Singapore, also known by its junior synonym, Duliticola hoiseni.
Platerodrilus is a genus of beetles of the family Lycidae. They commonly appear in the literature under the name Duliticola, which is an obsolete junior synonym.
The females retain a larval form as adults and are about 40–80 mm in length. The females and larvae have a flattened, dark body with large scales over the head, resembling trilobites, hence the informal names trilobite beetle, trilobite larva or Sumatran trilobite larva.
In contrast, the males are much smaller, 8–9 mm, resembling other beetles. Species are found in tropical rainforests of India and South-east Asia.
Trilobites ( meaning "three lobes") are a group of extinct marine artiopodan arthropods that form the
class Trilobita.
Trilobites form one of the earliest-known groups of arthropods. The first appearance of trilobites in the fossil record defines the base of the Atdabanian stage of the Early Cambrian period (521 million years ago), and they flourished throughout the lower Paleozoic before slipping into a long decline, when, during the Devonian, all trilobite orders except the Proetida died out. The last extant trilobites finally disappeared in the mass extinction at the end of the Permian about 252 million years ago. Trilobites were among the most successful of all early animals, existing in oceans for almost 300 million years.
By the time trilobites first appeared in the fossil record, they were already highly diversified and geographically dispersed. Because trilobites had wide diversity and an easily fossilized exoskeleton, they left an extensive fossil record. The study of their fossils has facilitated important contributions to biostratigraphy, paleontology, evolutionary biology, and plate tectonics.
Trilobites are often placed within the arthropod subphylum Schizoramia within the superclass Arachnomorpha (equivalent to the Arachnata), although several alternative taxonomies are found in the literature. More recently they have been placed within the Artiopoda, which includes many organisms that are morphologically similar to trilobites, but are largely unmineralised.
Trilobites had many lifestyles; some moved over the seabed as predators, scavengers, or filter feeders, and some swam, feeding on plankton. Some even crawled onto land.
Most lifestyles expected of modern marine arthropods are seen in trilobites, with the possible exception of parasitism (where scientific debate continues). Some trilobites (particularly the family Olenidae) are even thought to have evolved a symbiotic relationship with sulfur-eating bacteria from which they derived food.
The largest trilobites were more than 45 centimetres (18 in) long and may have weighed as much as 4.5 kilograms (9.9 lb).
Surprisingly enough, even the world's leading experts on these beetles are not 100% sure what they feed on, and up until relatively recently, there was some conflict as to what their diet consisted of.
However, the most popular and widely accepted theory is that these beetles feed on microorganisms living within the juices of rotten logs, as outlined in the findings of Alvin T. C. Wong (1996). Most Platerodrilus experts agree with Wong on that, so for now it should be treated as fact. Besides theres information that this sp do feed on slime mold too.
Kingdom: Animalia
Phylum: Arthropoda
Class: Insecta
Order: Coleoptera
Family: Lycidae
Genus: Platerodrilus
Species: P. ruficollis
Binomial name Platerodrilus ruficollis
(Pic, 1942)









Polydesmida
Flat back / Tractor millipede
(Barydesmus sp.)
Flat back / Tractor millipede
(Barydesmus sp.)


Millipedes are a group of arthropods that are characterised by having two pairs of jointed legs on most body segments; they are known scientifically as the class Diplopoda, the name being derived from this feature.
Each double-legged segment is a result of two single segments fused together. Most millipedes have very elongated cylindrical or flattened bodies with more than 20 segments, while pill millipedes are shorter and can roll into a ball.
Most millipedes are slow-moving detritivores, eating decaying leaves and other dead plant matter. Some eat fungi or suck plant fluids, and a small minority are predatory.
Millipedes are generally harmless to humans, although some can become household or garden pests. Millipedes can be unwanted especially in greenhouses where they can cause severe damage to emergent seedlings.
Most millipedes defend themselves with a variety of chemicals secreted from pores along the body, although the tiny bristle millipedes are covered with tufts of detachable bristles. Its primary defence mechanism is to curl into a tight coil, thereby protecting its legs and other vital delicate areas on the body behind a hard exoskeleton. Reproduction in most species is carried out by modified male legs called gonopods, which transfer packets of sperm to females.
First appearing in the Silurian period, millipedes are some of the oldest known land animals. Some members of prehistoric groups grew to over 2 m (6 ft 7 in); the largest modern species reach maximum lengths of 27 to 38 cm (11 to 15 in). The longest extant species is the giant African millipede (Archispirostreptus gigas).
Among myriapods, millipedes have traditionally been considered most closely related to the tiny pauropods, although some molecular studies challenge this relationship.
Millipedes can be distinguished from the somewhat similar but only distantly related centipedes (class Chilopoda), which move rapidly, are venomous, carnivorous, and have only a single pair of legs on each body segment.
The scientific study of millipedes is known as diplopodology, and a scientist who studies them is called a diplopodologist.
Polydesmida (from the Greek poly "many" and desmos "bond") is the largest order of millipedes, containing approximately 3,500 species, including all the millipedes reported to produce hydrogen cyanide (HCN).
Members of the order Polydesmida are also known as "flat-backed millipedes", because on most species, each body segment has wide lateral keels known as paranota. These keels are produced by the posterior half (metazonite) of each body ring behind the collum.
Polydesmids have no eyes, and vary in length from 3 to 130 mm (0.12 to 5.12 in). Including the telson, adults have 19 or 20 rings, while juveniles may have from 7 to 19 rings.
Mature males have a single pair of gonopods consisting of the modified forward leg pair of the 7th segment. Many of the larger species show bright coloration patterns which warn predators of their toxic secretions.
Polydesmids are very common in leaf litter, where they burrow by levering with the anterior end of the body.
The c. 3500 species of Polydesmida are variously classified into four suborders (names ending in "-idea"), and 29 families, the largest (numerically) including Paradoxosomatidae, Xystodesmidae, and Chelodesmidae.
Barydesmus sp is in the family Platyrhacidae found in tropical Asia and sourh America
Phylum: Arthropoda
Subphylum: Myriapoda
Class: Diplopoda
Superorder: Merocheta Cook, 1895
Order: Polydesmida Leach 1815
( flat-backed / tractor millipedes)
Genus: Barydesmus
Each double-legged segment is a result of two single segments fused together. Most millipedes have very elongated cylindrical or flattened bodies with more than 20 segments, while pill millipedes are shorter and can roll into a ball.
Most millipedes are slow-moving detritivores, eating decaying leaves and other dead plant matter. Some eat fungi or suck plant fluids, and a small minority are predatory.
Millipedes are generally harmless to humans, although some can become household or garden pests. Millipedes can be unwanted especially in greenhouses where they can cause severe damage to emergent seedlings.
Most millipedes defend themselves with a variety of chemicals secreted from pores along the body, although the tiny bristle millipedes are covered with tufts of detachable bristles. Its primary defence mechanism is to curl into a tight coil, thereby protecting its legs and other vital delicate areas on the body behind a hard exoskeleton. Reproduction in most species is carried out by modified male legs called gonopods, which transfer packets of sperm to females.
First appearing in the Silurian period, millipedes are some of the oldest known land animals. Some members of prehistoric groups grew to over 2 m (6 ft 7 in); the largest modern species reach maximum lengths of 27 to 38 cm (11 to 15 in). The longest extant species is the giant African millipede (Archispirostreptus gigas).
Among myriapods, millipedes have traditionally been considered most closely related to the tiny pauropods, although some molecular studies challenge this relationship.
Millipedes can be distinguished from the somewhat similar but only distantly related centipedes (class Chilopoda), which move rapidly, are venomous, carnivorous, and have only a single pair of legs on each body segment.
The scientific study of millipedes is known as diplopodology, and a scientist who studies them is called a diplopodologist.
Polydesmida (from the Greek poly "many" and desmos "bond") is the largest order of millipedes, containing approximately 3,500 species, including all the millipedes reported to produce hydrogen cyanide (HCN).
Members of the order Polydesmida are also known as "flat-backed millipedes", because on most species, each body segment has wide lateral keels known as paranota. These keels are produced by the posterior half (metazonite) of each body ring behind the collum.
Polydesmids have no eyes, and vary in length from 3 to 130 mm (0.12 to 5.12 in). Including the telson, adults have 19 or 20 rings, while juveniles may have from 7 to 19 rings.
Mature males have a single pair of gonopods consisting of the modified forward leg pair of the 7th segment. Many of the larger species show bright coloration patterns which warn predators of their toxic secretions.
Polydesmids are very common in leaf litter, where they burrow by levering with the anterior end of the body.
The c. 3500 species of Polydesmida are variously classified into four suborders (names ending in "-idea"), and 29 families, the largest (numerically) including Paradoxosomatidae, Xystodesmidae, and Chelodesmidae.
Barydesmus sp is in the family Platyrhacidae found in tropical Asia and sourh America
Phylum: Arthropoda
Subphylum: Myriapoda
Class: Diplopoda
Superorder: Merocheta Cook, 1895
Order: Polydesmida Leach 1815
( flat-backed / tractor millipedes)
Genus: Barydesmus



Bipalium kewense
Hammerhead flatworm
Hammerhead flatworm

Bipalium kewense is a species of large predatory land planarian with a cosmopolitan distribution. It is sometimes referred to as a "hammerhead flatworm" due to its half-moon-shaped head, but this name is also used to refer to other species in the subfamily Bipaliinae.
Bipalium kewense is a very long land planarian. Preserved specimens are up to 20 centimetres in length, and living specimens may be longer.
The anterior end ("head") is expanded in a transversal semilunate shape and the body is the narrowest just behind the head, in a region called "neck". The dorsal color is light-brown with five black to grey longitudinal stripes that begin at the neck. The median and marginal stripes are narrow and black, very distinctly marked. The lateral stripes (between the median and marginal stripes) are usually grey, broad and with diffuse margins. The neck is usually marked by an incomplete black collar formed by the union of the marginal and lateral stripes, this being one of the main characteristics to distinguish it from similar species. The head usually has a darker color than the background color of the dorsum and lacks stripes. The ventral side has a light ochre color externally and whitish over the creeping sole, which is externally lined by two diffuse grey-violet longitudinal lines.
Bipalium kewense is believed to be native to Southeast Asia, but currently is found worldwide. It was probably introduced by international plant trade, as it is frequently found associated with plant pots.
The species was first found in 1878 in the area of the Kew Park in the London Borough of Richmond upon Thames, United Kingdom, hence the name kewense.
B. kewense is a known predator of earthworms. It immobilizes the prey using muscular movements and possibly toxins and then everts its pharynx, connecting it to the earthworm's body and beginning digestion. The digestion seems to be at least partially extracorporeal by means of a collagenolyticenzyme.
Bipalium kewense is one of the few terrestrial invertebrates known to produce tetrodotoxin, a neurotoxin that results in paralysis. It is possible that the toxin aids the planarian in subduing its prey as well as in protecting it against predators.
All species of Bipalium are hermaphroditic, but Bipalium kewense has rarely been observed using sexual reproduction as a primary means of reproduction.
A sexual fragmentation is the primary means of reproduction in B. kewense in temperate regions. Long specimens usually release body fragments at the posterior end by transverse fission. The fragments are motile and regenerate the head plate and pharynx in a few weeks. Such a reproduction strategy is considered one of the reason for the successful colonization of this and other species of Bipalium.
Although there is little evidence of sexual reproduction in these planarians, there have been several reported cases of egg capsules being discovered. The egg capsules discovered had several of the same characteristics of those of B. adventitium, including coloration and incubation period. The most recent egg capsule discovered hatched offspring that did not bear a complete resemblance to adults and were considerably larger in size to that of B. adventitum offspring.
Order: Tricladida
Suborder: Continenticola
Family: Geoplanidae
Subfamily: Bipaliinae
Genus: Bipalium
Species: B. kewense
Binomial name Bipalium kewense
Moseley, 1878
Bipalium kewense is a very long land planarian. Preserved specimens are up to 20 centimetres in length, and living specimens may be longer.
The anterior end ("head") is expanded in a transversal semilunate shape and the body is the narrowest just behind the head, in a region called "neck". The dorsal color is light-brown with five black to grey longitudinal stripes that begin at the neck. The median and marginal stripes are narrow and black, very distinctly marked. The lateral stripes (between the median and marginal stripes) are usually grey, broad and with diffuse margins. The neck is usually marked by an incomplete black collar formed by the union of the marginal and lateral stripes, this being one of the main characteristics to distinguish it from similar species. The head usually has a darker color than the background color of the dorsum and lacks stripes. The ventral side has a light ochre color externally and whitish over the creeping sole, which is externally lined by two diffuse grey-violet longitudinal lines.
Bipalium kewense is believed to be native to Southeast Asia, but currently is found worldwide. It was probably introduced by international plant trade, as it is frequently found associated with plant pots.
The species was first found in 1878 in the area of the Kew Park in the London Borough of Richmond upon Thames, United Kingdom, hence the name kewense.
B. kewense is a known predator of earthworms. It immobilizes the prey using muscular movements and possibly toxins and then everts its pharynx, connecting it to the earthworm's body and beginning digestion. The digestion seems to be at least partially extracorporeal by means of a collagenolyticenzyme.
Bipalium kewense is one of the few terrestrial invertebrates known to produce tetrodotoxin, a neurotoxin that results in paralysis. It is possible that the toxin aids the planarian in subduing its prey as well as in protecting it against predators.
All species of Bipalium are hermaphroditic, but Bipalium kewense has rarely been observed using sexual reproduction as a primary means of reproduction.
A sexual fragmentation is the primary means of reproduction in B. kewense in temperate regions. Long specimens usually release body fragments at the posterior end by transverse fission. The fragments are motile and regenerate the head plate and pharynx in a few weeks. Such a reproduction strategy is considered one of the reason for the successful colonization of this and other species of Bipalium.
Although there is little evidence of sexual reproduction in these planarians, there have been several reported cases of egg capsules being discovered. The egg capsules discovered had several of the same characteristics of those of B. adventitium, including coloration and incubation period. The most recent egg capsule discovered hatched offspring that did not bear a complete resemblance to adults and were considerably larger in size to that of B. adventitum offspring.
Order: Tricladida
Suborder: Continenticola
Family: Geoplanidae
Subfamily: Bipaliinae
Genus: Bipalium
Species: B. kewense
Binomial name Bipalium kewense
Moseley, 1878
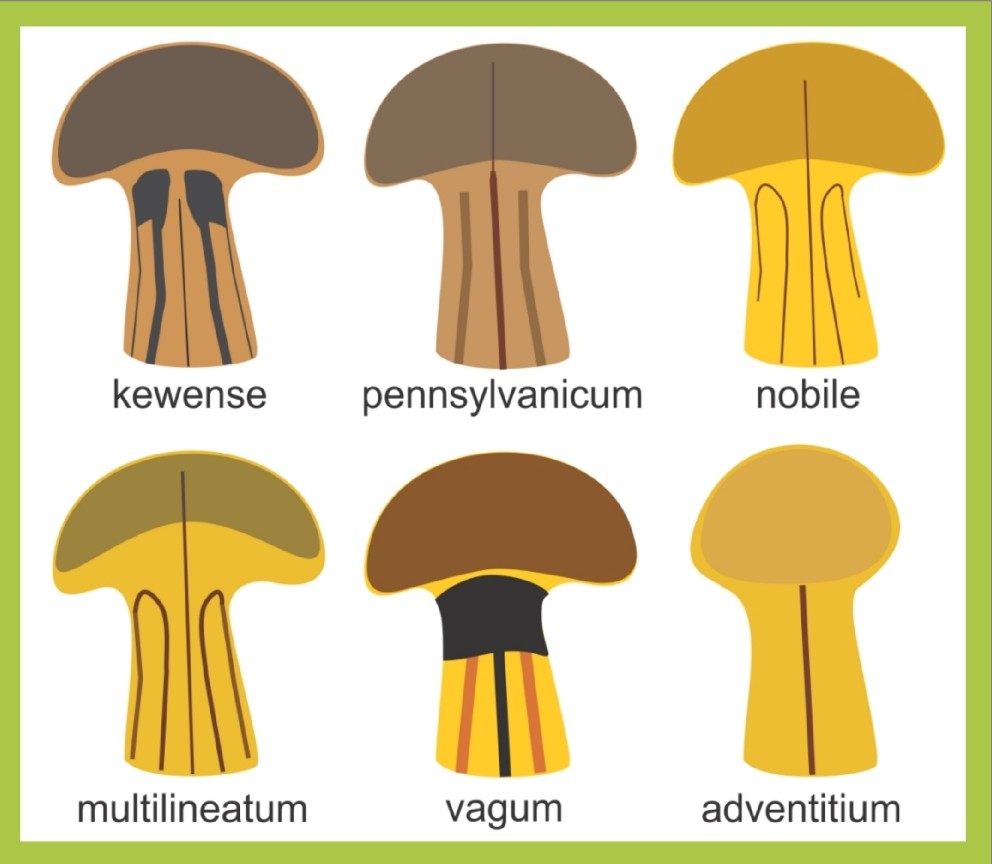

Sphaerotheriida
giant pill millipedes.
giant pill millipedes.

Pill millipedes are any members of two living (and one extinct) orders of millipedes, often grouped together into a single superorder, Oniscomorpha. The name Oniscomorpha refers to the millipedes' resemblance to certain woodlice (Oniscidea), also called pillbugs or "roly-polies". However, millipedes and woodlice are not closely related (belonging to the subphyla Myriapoda and Crustacea, respectively); rather, this is a case of convergent evolution.
Pill millipedes are relatively short-bodied compared to most other millipedes, with only eleven to thirteen body segments, and are capable of rolling into a ball (volvation) when disturbed, as a defense against predators. This ability evolved separately in each of the two orders, making it a case of convergent evolution, rather than homology. They can also exude a noxious liquid, which may be both caustic and toxic, to repel predators. Pill millipedes are detritivorous, feeding on decomposing plant matter, usually in woodlands.
The order Glomerida is predominantly found in the Northern Hemisphere and includes species such as Glomeris marginata, the common European pill millipede. They have from eleven to twelve body segments, and possess dorsal ozopores (openings of the repugnatorial glands) rather than the lateral ozopores found on many other millipedes. Glomeridans reach maximum lengths of 20 mm (0.79 in), and eyes, if present, are in a single row of ocelli.
The order contains approximately 450 species found in Europe, South-east Asia and the Americas from California to Guatemala. Four species are present in the British Isles.
The order Sphaerotheriida is a Gondwana-distribution taxon, with around 350 species in southern Africa, Madagascar, Australasia and South East Asia. Five species, all in the genus Procyliosoma are present in New Zealand and around thirty species are present in Australia. Sphaerotheriidans have thirteen body segments, and do not possess repugnatorial glands. Spherotheriidans reach larger size than Glomeridans (up to 10 cm (3.9 in)), and always possess large, kidney-shaped eyes.
Sphaerotheriida is an order of millipedes in the infraclass Pentazonia, sometimes known as giant pill millipedes.
They inhabit Southern Africa, Madagascar, South and Southeast Asia, Australia and New Zealand.
Like the Northern Hemisphere pill millipedes of the order Glomerida, these millipedes can roll into a ball when disturbed. When they are rolled-up, most sphaerotheriidans reach a maximum size of a cherry or golf ball, but some species from Madagascar can even reach the size of an orange.
When rolled-up, predators are unable to unravel giant pill millipedes since the margins of their second and last dorsal plates fit perfectly into one another, creating a sealed ball. A few giant pill millipede species are able to produce sound, the only millipedes known to do this. This order of millipedes is also unique in that some African species are used for medicinal purposes.
Sphaerotheriidans are characterized by a relatively conservative body morphology; superficially all species and genera look the same.
Dorsally, their head is followed by twelve body tergites (collum, thoracic shield, and ten normal tergites) and the anal shield.
Ventrally, females possess twenty-one leg pairs (forty-two legs in total), while males carry two additional modified leg pairs, the anterior and posterior telopods under their anal shield. The telopods resemble chelae and/or clamp-like structures, which are probably used in holding the female during mating. In the position of the male telopods, the females instead have a sclerotized subanal plate, which in some species such as those belonging to the family Arthrosphaeridae, is enlarged and is used to produce vibrations (stridulation). Furthermore, unlike other large-bodied millipede orders, Sphaerotheriida do not have glands that excrete poisonous or ill-smelling substances. Instead they depend entirely on their rolling-up behavior for protection.
Sphaerotheriida somewhat resemble the North American and Eurasian pill millipedes of the order Glomerida, but are generally larger in size (20–80 millimetres or 0.8–3.1 inches body length). Until the end of the 20th century only the largest known species in the order Glomerida rivalled the size of even the smallest known Sphaerotheriida, but in the early 21st century a much smaller Sphaerotheriid was described from Madagascar: full-grown specimens of Microsphaerotherium ivohibiensis are just the size of a pea.
Also on Madagascar, some giant pill millipede species exhibit island gigantism, reaching more than 9.5 cm (3.7 in) in outstretched length and a size comparable to an orange when rolled up.
The orders differ in the number of tergites (10 or 11 in Glomerida, 12 in Sphaerotheriida) and legs (17 or 19 in Glomerida, 21 or 23 in Sphaerotheriida), and show great differences in their head morphology and genital openings, among other traits.
Both orders have the ability to roll into a perfect ball, protecting the head, antennae, and the vulnerable underside. However, this rolled-up position (volvation) is achieved differently.
In Glomerida, the enlarged second body ring (thoracic shield) has a more or less visible gap within which fit the tips of tergites 3–11, whereas in Sphaerotheriida the tips of tergites 3–12 fit perfectly into a groove on the thoracic shield. Juvenile sphaerotheriidans show the same gap as the Glomerida.
Many giant pill millipede species have special ledges ('locking carinae') on the underside of the tergite tips and the anal shield which can be moved above a brim on the thoracic shield. These millipedes remain passively locked-up since they need not continued muscle contraction to remain in the rolled-up position.
In general, Sphaerotheriida have a Gondwanan distribution.
Little is known about the ecology, development and life history of Sphaerotheriida, but apparently all species are detritivores, feeding on dead organic matter such as leaves and wood on the forest floor. Like earthworms, they play an important role in decomposition; by breaking down decaying organic matter they release locked up nutrients back into the soil. Such recycling is essential for plant nutrition and accordingly for the entire ecology.
It is possible that giant pill millipedes rely on special bacteria in their gut, much as termites do, to exploit the nutritional value of otherwise indigestible material such as lignin.
Sphaerotheriida inhabit mainly the leaf litter of humid forests. Some species, however, show an arboreal (tree-living) lifestyle, and in these the rolling-up reflex has been suppressed.
The rolling-up ability and tough skeletal armor of the Sphaerotheriida offer protection against some predators, but a wide variety of predators feed on them, or even specialise in them as a source of food.
Species that specialize in feeding on giant pill millipedes necessarily have evolved special structures or behaviors to overcome their defences.
Examples include the South African snail family Chlamydephoridae which almost exclusively feeds on giant pill millipedes.
Another example is the meerkat (Suricata suricata) which has been reported (at least in captivity) to throw rolled-up sphaerotheriids against rocks in order to break them. This behaviour however, is their way of breaking open many refractory food items, such as snails and hard-shelled eggs.
Apart from the rolling-up behavior of giant pill millipedes, camouflage may be an important defense mechanism against predators that hunt by sight, such as birds.
Sphaerotheriida also are subject to internal parasitism, and several species of nematodes are obligate parasites of particular species of giant pill millipedes.
Phylum: Arthropoda
Class: Diplopoda
Subclass: Chilognatha
Infraclass: Pentazonia
Superorder: Oniscomorpha
Order: Sphaerotheriida
Brandt, 1833Families
Pill millipedes are relatively short-bodied compared to most other millipedes, with only eleven to thirteen body segments, and are capable of rolling into a ball (volvation) when disturbed, as a defense against predators. This ability evolved separately in each of the two orders, making it a case of convergent evolution, rather than homology. They can also exude a noxious liquid, which may be both caustic and toxic, to repel predators. Pill millipedes are detritivorous, feeding on decomposing plant matter, usually in woodlands.
The order Glomerida is predominantly found in the Northern Hemisphere and includes species such as Glomeris marginata, the common European pill millipede. They have from eleven to twelve body segments, and possess dorsal ozopores (openings of the repugnatorial glands) rather than the lateral ozopores found on many other millipedes. Glomeridans reach maximum lengths of 20 mm (0.79 in), and eyes, if present, are in a single row of ocelli.
The order contains approximately 450 species found in Europe, South-east Asia and the Americas from California to Guatemala. Four species are present in the British Isles.
The order Sphaerotheriida is a Gondwana-distribution taxon, with around 350 species in southern Africa, Madagascar, Australasia and South East Asia. Five species, all in the genus Procyliosoma are present in New Zealand and around thirty species are present in Australia. Sphaerotheriidans have thirteen body segments, and do not possess repugnatorial glands. Spherotheriidans reach larger size than Glomeridans (up to 10 cm (3.9 in)), and always possess large, kidney-shaped eyes.
Sphaerotheriida is an order of millipedes in the infraclass Pentazonia, sometimes known as giant pill millipedes.
They inhabit Southern Africa, Madagascar, South and Southeast Asia, Australia and New Zealand.
Like the Northern Hemisphere pill millipedes of the order Glomerida, these millipedes can roll into a ball when disturbed. When they are rolled-up, most sphaerotheriidans reach a maximum size of a cherry or golf ball, but some species from Madagascar can even reach the size of an orange.
When rolled-up, predators are unable to unravel giant pill millipedes since the margins of their second and last dorsal plates fit perfectly into one another, creating a sealed ball. A few giant pill millipede species are able to produce sound, the only millipedes known to do this. This order of millipedes is also unique in that some African species are used for medicinal purposes.
Sphaerotheriidans are characterized by a relatively conservative body morphology; superficially all species and genera look the same.
Dorsally, their head is followed by twelve body tergites (collum, thoracic shield, and ten normal tergites) and the anal shield.
Ventrally, females possess twenty-one leg pairs (forty-two legs in total), while males carry two additional modified leg pairs, the anterior and posterior telopods under their anal shield. The telopods resemble chelae and/or clamp-like structures, which are probably used in holding the female during mating. In the position of the male telopods, the females instead have a sclerotized subanal plate, which in some species such as those belonging to the family Arthrosphaeridae, is enlarged and is used to produce vibrations (stridulation). Furthermore, unlike other large-bodied millipede orders, Sphaerotheriida do not have glands that excrete poisonous or ill-smelling substances. Instead they depend entirely on their rolling-up behavior for protection.
Sphaerotheriida somewhat resemble the North American and Eurasian pill millipedes of the order Glomerida, but are generally larger in size (20–80 millimetres or 0.8–3.1 inches body length). Until the end of the 20th century only the largest known species in the order Glomerida rivalled the size of even the smallest known Sphaerotheriida, but in the early 21st century a much smaller Sphaerotheriid was described from Madagascar: full-grown specimens of Microsphaerotherium ivohibiensis are just the size of a pea.
Also on Madagascar, some giant pill millipede species exhibit island gigantism, reaching more than 9.5 cm (3.7 in) in outstretched length and a size comparable to an orange when rolled up.
The orders differ in the number of tergites (10 or 11 in Glomerida, 12 in Sphaerotheriida) and legs (17 or 19 in Glomerida, 21 or 23 in Sphaerotheriida), and show great differences in their head morphology and genital openings, among other traits.
Both orders have the ability to roll into a perfect ball, protecting the head, antennae, and the vulnerable underside. However, this rolled-up position (volvation) is achieved differently.
In Glomerida, the enlarged second body ring (thoracic shield) has a more or less visible gap within which fit the tips of tergites 3–11, whereas in Sphaerotheriida the tips of tergites 3–12 fit perfectly into a groove on the thoracic shield. Juvenile sphaerotheriidans show the same gap as the Glomerida.
Many giant pill millipede species have special ledges ('locking carinae') on the underside of the tergite tips and the anal shield which can be moved above a brim on the thoracic shield. These millipedes remain passively locked-up since they need not continued muscle contraction to remain in the rolled-up position.
In general, Sphaerotheriida have a Gondwanan distribution.
Little is known about the ecology, development and life history of Sphaerotheriida, but apparently all species are detritivores, feeding on dead organic matter such as leaves and wood on the forest floor. Like earthworms, they play an important role in decomposition; by breaking down decaying organic matter they release locked up nutrients back into the soil. Such recycling is essential for plant nutrition and accordingly for the entire ecology.
It is possible that giant pill millipedes rely on special bacteria in their gut, much as termites do, to exploit the nutritional value of otherwise indigestible material such as lignin.
Sphaerotheriida inhabit mainly the leaf litter of humid forests. Some species, however, show an arboreal (tree-living) lifestyle, and in these the rolling-up reflex has been suppressed.
The rolling-up ability and tough skeletal armor of the Sphaerotheriida offer protection against some predators, but a wide variety of predators feed on them, or even specialise in them as a source of food.
Species that specialize in feeding on giant pill millipedes necessarily have evolved special structures or behaviors to overcome their defences.
Examples include the South African snail family Chlamydephoridae which almost exclusively feeds on giant pill millipedes.
Another example is the meerkat (Suricata suricata) which has been reported (at least in captivity) to throw rolled-up sphaerotheriids against rocks in order to break them. This behaviour however, is their way of breaking open many refractory food items, such as snails and hard-shelled eggs.
Apart from the rolling-up behavior of giant pill millipedes, camouflage may be an important defense mechanism against predators that hunt by sight, such as birds.
Sphaerotheriida also are subject to internal parasitism, and several species of nematodes are obligate parasites of particular species of giant pill millipedes.
Phylum: Arthropoda
Class: Diplopoda
Subclass: Chilognatha
Infraclass: Pentazonia
Superorder: Oniscomorpha
Order: Sphaerotheriida
Brandt, 1833Families


Harpaphe haydeniana
Yellow-Spotted Millipede
Yellow-Spotted Millipede

Harpaphe haydeniana, commonly known as the yellow-spotted millipede, almond-scented millipede or cyanide millipede, is a species of polydesmidan ("flat-backed") millipede found in the moist forests along the Pacific coast of North America, from Southeast Alaska to California.
The dark coloration with contrasting yellow-tipped keels warn of its ability to exude toxic hydrogen cyanide as a defense.
Despite the various common names given the species, the coloration pattern, cyanide defense, and associated almond scent occur in other flat-backed millipedes around the world.
Phylum: Arthropoda
Class: Diplopoda
Order: Polydesmida
Family: Xystodesmidae
Genus: Harpaphe
Species: H. haydeniana
Binomial name Harpaphe haydeniana
The dark coloration with contrasting yellow-tipped keels warn of its ability to exude toxic hydrogen cyanide as a defense.
Despite the various common names given the species, the coloration pattern, cyanide defense, and associated almond scent occur in other flat-backed millipedes around the world.
Phylum: Arthropoda
Class: Diplopoda
Order: Polydesmida
Family: Xystodesmidae
Genus: Harpaphe
Species: H. haydeniana
Binomial name Harpaphe haydeniana

Trigoniulus corallinus
Rusty Millipede
Rusty Millipede

Trigoniulus corallinus, sometimes called the rusty millipede or common Asian millipede, is a species of millipede widely distributed in the Indo-Malayan region including India, Sri Lanka, China, Taiwan, Myanmar, Thailand, Vietnam, Malaysia, Singapore, and much of Indonesia.
It is also reported from Fiji and Tanzania and found in South Asia and the Caribbean as an introduced species.
These millipedes inhabit moist areas, rotten wood and compost. The genome of T. corallinus was sequenced in 2015, the first time this has been done for a millipede.
Phylum: Arthropoda
Class: Diplopoda
Order: Spirobolida
Family: Trigoniulidae
Genus: Trigoniulus
Species: T. corallinus
Binomial name Trigoniulus corallinus
(Gervais, 1842)
It is also reported from Fiji and Tanzania and found in South Asia and the Caribbean as an introduced species.
These millipedes inhabit moist areas, rotten wood and compost. The genome of T. corallinus was sequenced in 2015, the first time this has been done for a millipede.
Phylum: Arthropoda
Class: Diplopoda
Order: Spirobolida
Family: Trigoniulidae
Genus: Trigoniulus
Species: T. corallinus
Binomial name Trigoniulus corallinus
(Gervais, 1842)

Scutigera coleoptrata
House Centipede
House Centipede

Scutigera coleoptrata is a small, typically yellowish-grey centipede with up to 15 pairs of long legs. Originating in the Mediterranean region, the species has spread to other parts of the world, where it can live in human homes, thus gaining the name house centipede. It is an insectivore; it kills and eats other arthropods, such as insects and arachnids.
Two of the house centipede’s legs, located right near the head and near the mouth, have been modified to carry venom. Technically, this means that house centipede sting their prey rather than bite. Their venom is potent for smaller insects such as silverfish and termites. They are also capable of holding multiple preys in their legs and if they get any of their legs caught, they can just break them off and scurry away. Their eyes are very sensitive to light, so more than likely they will find a place to hide during the day.
They are not considered dangerous to humans. While it is possible that one might bite a person, more than likely it would take picking up a house centipede and handling one for that to happen. They would much rather reserve their venom for food and humans just are not something on the menu.
If someone is bitten, more than likely it will just cause a reddish bump. People who are especially sensitive to bee stings and other insect bites and stings may need to see a doctor to make sure they don’t have an allergic reaction to the house centipede bite, but most people should feel no effects at all.
The body of an adult Scutigera coleoptrata is 25 to 35 mm (0.98 to 1.38 in) in length. Up to 15 pairs of long legs are attached to the rigid body. Together with the antennae they give the centipede an appearance of being 75 to 100 mm (3 to 4 in) in length. The delicate legs enable it to reach surprising speeds of up to 0.4 meters per second (1.3 ft/s) running across floors, up walls and along ceilings. Its body is yellowish-grey and has three dark dorsal stripes running down its length; the legs also have dark stripes. S. coleoptrata has developed automimicry in that its tail-like hind legs present the appearance of antennae. When the centipede is at rest, it is not easy to tell its cranial end from its caudal end.
Unlike most other centipedes, house centipedes and their close relatives have well-developed faceted eyes.
House centipedes live anywhere from three to seven years, depending on the environment. They can start breeding in their third year. To begin mating, the male and female circle around each other. They initiate contact with their antennae. The male deposits his sperm on the ground and the female then uses it to fertilize her eggs.
House centipedes feed on spiders, bed bugs, termites, cockroaches, silverfish, ants, and other household arthropods. They administer venom through forcipules. These are not part of their mandibles, so strictly speaking they sting rather than bite. They are mostly nocturnal hunters. Despite their developed eyes, they seem to rely mostly on their antennae when hunting. Their antennae are sensitive to both smells and tactile information. They use both their mandibles and their legs for holding prey. This way they can deal with several small insects at the same time. To capture prey they either jump onto it or use their legs in a technique described as "lassoing". Using their legs to beat prey has also been described. Like other centipedes they can stridulate.
Outdoors, house centipedes prefer to live in cool, damp places. Centipede respiratory systems do not provide any mechanism for shutting the spiracles, and that is why they need an environment that protects them from dehydration and excessive cold. Most live outside, primarily under large rocks, piles of wood, and especially in compost piles. Within the home, these centipedes are found in almost any part of the house. Most commonly they are encountered in basements, bathrooms, and lavatories, which tend to be humid, but they can also be found in drier places like offices, bedrooms and dining rooms. The greatest likelihood of encountering them is in spring, when they come out because the weather gets warmer, and in autumn/fall, when the cooling weather forces them to find shelter in human habitats.
The faceted eyes of S. coleoptrata are sensitive to daylight and very sensitive to ultraviolet light. They were shown to be able to visually distinguish between different mutations of Drosophila melanogaster.
How this ability fits with its nocturnal lifestyle and underground natural habitat is still under study. They do not instantly change direction when light is suddenly shone at them, but will retreat to a darker hiding spot.
Some of the plates covering the body segments fused and became smaller during the evolution to the current state of S. coleoptrata. The resulting mismatch between body segments and dorsal plates (tergites) is the cause for this centipede's rigid body.
Unlike its shorter-legged but much larger tropical cousins, S. coleoptrata can live its entire life inside a building, usually the ground levels of homes. Though they often startle their unwitting housemates with their appearance and surprising speed, they are generally considered harmless to humans. Bites are uncommon, and the forcipules of house centipedes are not strong enough to easily penetrate human skin. Its bite and venom are typically less severe than other centipedes.
Techniques for eliminating centipedes from homes include drying up the areas where they thrive, eliminating large indoor insect populations, and sealing cracks in the walls. An alternative is to let them live, allowing them to control pests, e.g., roaches, flies, moths, bed bugs, silverfish, earwigs, etc.
Phylum: Arthropoda
Class: Chilopoda
Order: Scutigeromorpha
Family: Scutigeridae
Genus: Scutigera
Species: S. coleoptrata
Binomial name Scutigera coleoptrata
(Linnaeus, 1758)
Two of the house centipede’s legs, located right near the head and near the mouth, have been modified to carry venom. Technically, this means that house centipede sting their prey rather than bite. Their venom is potent for smaller insects such as silverfish and termites. They are also capable of holding multiple preys in their legs and if they get any of their legs caught, they can just break them off and scurry away. Their eyes are very sensitive to light, so more than likely they will find a place to hide during the day.
They are not considered dangerous to humans. While it is possible that one might bite a person, more than likely it would take picking up a house centipede and handling one for that to happen. They would much rather reserve their venom for food and humans just are not something on the menu.
If someone is bitten, more than likely it will just cause a reddish bump. People who are especially sensitive to bee stings and other insect bites and stings may need to see a doctor to make sure they don’t have an allergic reaction to the house centipede bite, but most people should feel no effects at all.
The body of an adult Scutigera coleoptrata is 25 to 35 mm (0.98 to 1.38 in) in length. Up to 15 pairs of long legs are attached to the rigid body. Together with the antennae they give the centipede an appearance of being 75 to 100 mm (3 to 4 in) in length. The delicate legs enable it to reach surprising speeds of up to 0.4 meters per second (1.3 ft/s) running across floors, up walls and along ceilings. Its body is yellowish-grey and has three dark dorsal stripes running down its length; the legs also have dark stripes. S. coleoptrata has developed automimicry in that its tail-like hind legs present the appearance of antennae. When the centipede is at rest, it is not easy to tell its cranial end from its caudal end.
Unlike most other centipedes, house centipedes and their close relatives have well-developed faceted eyes.
House centipedes live anywhere from three to seven years, depending on the environment. They can start breeding in their third year. To begin mating, the male and female circle around each other. They initiate contact with their antennae. The male deposits his sperm on the ground and the female then uses it to fertilize her eggs.
House centipedes feed on spiders, bed bugs, termites, cockroaches, silverfish, ants, and other household arthropods. They administer venom through forcipules. These are not part of their mandibles, so strictly speaking they sting rather than bite. They are mostly nocturnal hunters. Despite their developed eyes, they seem to rely mostly on their antennae when hunting. Their antennae are sensitive to both smells and tactile information. They use both their mandibles and their legs for holding prey. This way they can deal with several small insects at the same time. To capture prey they either jump onto it or use their legs in a technique described as "lassoing". Using their legs to beat prey has also been described. Like other centipedes they can stridulate.
Outdoors, house centipedes prefer to live in cool, damp places. Centipede respiratory systems do not provide any mechanism for shutting the spiracles, and that is why they need an environment that protects them from dehydration and excessive cold. Most live outside, primarily under large rocks, piles of wood, and especially in compost piles. Within the home, these centipedes are found in almost any part of the house. Most commonly they are encountered in basements, bathrooms, and lavatories, which tend to be humid, but they can also be found in drier places like offices, bedrooms and dining rooms. The greatest likelihood of encountering them is in spring, when they come out because the weather gets warmer, and in autumn/fall, when the cooling weather forces them to find shelter in human habitats.
The faceted eyes of S. coleoptrata are sensitive to daylight and very sensitive to ultraviolet light. They were shown to be able to visually distinguish between different mutations of Drosophila melanogaster.
How this ability fits with its nocturnal lifestyle and underground natural habitat is still under study. They do not instantly change direction when light is suddenly shone at them, but will retreat to a darker hiding spot.
Some of the plates covering the body segments fused and became smaller during the evolution to the current state of S. coleoptrata. The resulting mismatch between body segments and dorsal plates (tergites) is the cause for this centipede's rigid body.
Unlike its shorter-legged but much larger tropical cousins, S. coleoptrata can live its entire life inside a building, usually the ground levels of homes. Though they often startle their unwitting housemates with their appearance and surprising speed, they are generally considered harmless to humans. Bites are uncommon, and the forcipules of house centipedes are not strong enough to easily penetrate human skin. Its bite and venom are typically less severe than other centipedes.
Techniques for eliminating centipedes from homes include drying up the areas where they thrive, eliminating large indoor insect populations, and sealing cracks in the walls. An alternative is to let them live, allowing them to control pests, e.g., roaches, flies, moths, bed bugs, silverfish, earwigs, etc.
Phylum: Arthropoda
Class: Chilopoda
Order: Scutigeromorpha
Family: Scutigeridae
Genus: Scutigera
Species: S. coleoptrata
Binomial name Scutigera coleoptrata
(Linnaeus, 1758)




