Special Note...
The information in this fungus page
has been compiled from reliable sources through other webpage as for self reference learning works.
It is not a substitute
for food and or for any medicinal purposes and it does not purport to provide any advice.
In the field of wild fungus
each and every individual body absorbing system is different and It should not be consume or used without any expert guidance.
Readers should always consult the expert
in the field before consuming any wild fungus and for using it as for medicinal purposes.
The information in this fungus page
has been compiled from reliable sources through other webpage as for self reference learning works.
It is not a substitute
for food and or for any medicinal purposes and it does not purport to provide any advice.
In the field of wild fungus
each and every individual body absorbing system is different and It should not be consume or used without any expert guidance.
Readers should always consult the expert
in the field before consuming any wild fungus and for using it as for medicinal purposes.
Phallaceae induaiatus
sinkhorn
sinkhorn

Phallaceae is a family of fungi, commonly known as stinkhorns, within the order Phallales. Stinkhorns have a worldwide distribution, but are especially prevalent in tropical regions.
They are known for their foul-smelling, sticky spore
masses, or gleba, borne on the end of a stalk called the receptaculum. The characteristic fruiting-body structure, a single, unbranched receptaculum with an externally attached gleba on the upper part, distinguishes the Phallaceae from other families in the Phallales.
The spore mass typically smells of carrion or dung, and attracts flies, beetles and other insects to help disperse the spores. Although there is great diversity in body structure shape among the various genera, all species in the Phallaceae begin their development as oval or round structures known as "eggs".
The appearance of Phallaceae is often sudden, as gleba can erupt from the underground egg and burst open within an hour. According to a 2008 estimate, the family contains 21 genera and 77 species.
Species of stinkhorns have gasteroid, or internally produced spores. Fruit bodies originate as a gelatinous, spherical, or egg-shaped structure that may be completely or partially buried underground.
The peridium, the outer layer of the egg, is white, or purple/red, with two or three layers. The outer layer is thin, membranous, and elastic, while the inner layer is thicker, gelatinous, and continuous. At maturity, the peridium opens up and remains as a volva at the base of the receptaculum.
The fertile portion of the fruiting body is often borne on the end of a wide, fleshy or spongy stalk (as in the Phallales), which may be cylindrical, star-shaped, or reticulate (forming a network).
They may be brightly colored, sometimes with a lattice or veil-like membrane enclosing and protecting the spores. The spore-containing substance, the gleba, is typically gelatinous, often foul-smelling, and deliquescent (becoming liquid from the absorption of water). The gleba is formed on the exterior face of the cap or the upper part of the receptacle.
The basidia are small and narrowly club-shaped or fusiform, short-lived (evanescent), with four to eight sterigmata. The spores are usually ellipsoid or cylindrical in shape, hyaline or pale brown, smooth, more or less smooth-walled, and truncated at the base.
Phallus indusiatus, commonly called the bamboo mushrooms, bamboo pith, long net stinkhorn, crinoline stinkhorn or veiled lady, is a fungus in the family Phallaceae, or stinkhorns.
It has a cosmopolitan distribution in tropical areas, and is found in southern Asia, Africa, the Americas, and Australia, where it grows in woodlands and gardens in rich soil and well-rotted woody material.
The fruit body of the fungus is characterised by a conical to bell-shaped cap on a stalk and a delicate lacy "skirt", or indusium, that hangs from beneath the cap and reaches nearly to the ground. First described scientifically in 1798 by French botanist Étienne Pierre Ventenat, the species has often been referred to a separate genus Dictyophora along with other Phallus
\\\\nspecies featuring an indusium.
P. indusiatus can be distinguished from other similar species by differences in distribution, size, color, and indusium length. Mature fruit bodies are up to 25 cm (10 in) tall with a conical to bell-shaped cap that is 1.5–4 cm (0.6–1.6 in) wide. The cap is covered with a greenish-brown spore-containing slime, which attracts flies and other insects that eat the spores and disperse them.
An edible mushroom featured as an ingredient in Chinese haute cuisine, it is used in stir-fries and chicken soups. The mushroom, grown commercially and commonly sold in Asian markets, is rich in protein, carbohydrates, and dietary fiber. The mushroom also contains various bioactive compounds, and has antioxidant and antimicrobial properties.
Phallus indusiatus has a recorded history of use in Chinese medicine extending back to the 7th century AD, and features in Nigerian folklore.
Phallus indusiatus has many common names based on its appearance, including long net stinkhorn, crinoline stinkhorn, basket stinkhorn, bridal veil fungus, and veiled lady. A Chinese common name that alludes to its typical growth habitat is "bamboo mushroom" (simplified Chinese: 竹荪; traditional Chinese: 竹蓀; pinyin: zhúsūn).
Immature fruit bodies of P. indusiatus are initially enclosed in an egg-shaped to roughly spherical subterranean structure encased in a peridium.
The "egg" ranges in color from whitish to buff to reddish-brown, measures up to 6 cm (2.4 in) in diameter, and usually has a thick mycelial cord attached at the bottom. As the mushroom matures, the pressure caused by the enlargement of the internal structures cause the peridium to tear and the fruit body rapidly emerges from the "egg". The mature mushroom is up to 25 cm (9.8 in) tall and girded with a net-like structure called the indusium (or less technically a "skirt") that hangs down from the conical to bell-shaped cap.
The netlike openings of the indusium may be polygonal or round in shape. Well-developed specimens have an indusium that reaches to the volva and flares out somewhat before collapsing on the stalk.
The cap is 1.5–4 cm (0.6–1.6 in) wide and its reticulated (pitted and ridged) surface is covered with a layer of greenish-brown and foul-smelling slime, the gleba, which initially partially obscures the reticulations.
The top of the cap has a small hole. The stalk is 7–25 cm (2.8–9.8 in) long and 1.5–3 cm (0.6–1.2 in) thick. The hollow stalk is white, roughly equal in width throughout its length, sometimes curved, and spongy.
The ruptured peridium remains as a loose volva at the base of the stalk. Fruit bodies develop during the night and require 10–15 hours to fully develop after emerging from the peridium. They are short-lived, typically lasting no more than a few days. At that point the slime has usually been removed by insects, leaving the pale off-white, bare cap surface exposed.
Spores of P. indusiatus are thin-walled, smooth, elliptical or slightly curved, hyaline (translucent), and measure 2–3 by 1–1.5 μm.
Phallus multicolor is similar in overall appearance, but it has a more brightly coloured cap, stem and indusium, and it is usually smaller. It is found in Australia, Guam, Sumatra, Java, Borneo, Papua New Guinea, Zaire, and Tobago as well as Hawaii.
In contrast, the cap surface of P. indusiatus tends to have conspicuous reticulations that remain clearly visible under the gleba.
P. indusiatus is saprobic deriving nutrients from breaking down wood and plant organic matter. The fruit bodies grow singly or in groups in disturbed ground and among wood chips.
In eastern Asia, P. indusiatus is considered a delicacy and an aphrodisiac. Previously only collected in the wild, where it is not abundant, it was difficult to procure.
Phallus indusiatus has been cultivated on a commercial scale in China since 1979. In the Fujian Province of China known for a thriving mushroom industry that cultivates 45 species of edible fungi, P. indusiatus is produced in the counties of Fuan, Jianou, and Ningde.
The optimal temperature for the growth of mushroom spawn and fruit bodies is about 24 °C (75 °F), with a relative humidity of 90–95%. Other substrates that can be used for the cultivation of the fungus include bamboo leaves and small stems, soybean pods or stems, corn stems, and willow leaves.
Nutritional analyses of P. indusiatus show that the fruit bodies are over 90% water, about 6% fiber, 4.8% protein, 4.7% fat, and several mineral elements, including calcium, although the mineral composition in the fungus may depend on corresponding concentrations in the growth substrate.
Medicinal properties have been ascribed to Phallus indusiatus from the time of the Chinese Tang Dynasty when it was described in pharmacopoeia.
The fungus was used to treat many inflammatory,
stomach, and neural diseases. Southern China's Miao people continue to use it traditionally for a number of afflictions, including injuries and pains, cough,
dysentery, enteritis, leukemia, and feebleness, and it has been prescribed clinically as a treatment for laryngitis, leucorrhea, fever, and oliguria (low urine output), diarrhea, hypertension, cough, hyperlipidemia, and in anticancer therapy. Modern science has probed the biochemical basis of these putative medicinal benefits.
The fungus has long been recognised to have
antibacterial properties: the addition of the fungus to soup broth was known to prevent it from spoiling for several days.
One of the responsible antibiotics, albaflavenone, was isolated in 2011. It is a sesquiterpenoid that was already known from the soil bacterium Streptomyces albidoflavus. Experiments have shown that extracts of P. indusiatus have antioxidant in addition to antimicrobial properties.
Kingdom : Fungi
Division : Basidiomycota
Class :vAgaricomycetes
Order : Phallales
Family : Phallaceae
Genus : Phallus
Species : P. indusiatus
Binomial name Phallus indusiatus
Vent. (1798)
Synonyms :
Dictyophora indusiata (Vent.) Desv. (1809)
Hymenophallus indusiatus (Vent.) Nees (1817)
They are known for their foul-smelling, sticky spore
masses, or gleba, borne on the end of a stalk called the receptaculum. The characteristic fruiting-body structure, a single, unbranched receptaculum with an externally attached gleba on the upper part, distinguishes the Phallaceae from other families in the Phallales.
The spore mass typically smells of carrion or dung, and attracts flies, beetles and other insects to help disperse the spores. Although there is great diversity in body structure shape among the various genera, all species in the Phallaceae begin their development as oval or round structures known as "eggs".
The appearance of Phallaceae is often sudden, as gleba can erupt from the underground egg and burst open within an hour. According to a 2008 estimate, the family contains 21 genera and 77 species.
Species of stinkhorns have gasteroid, or internally produced spores. Fruit bodies originate as a gelatinous, spherical, or egg-shaped structure that may be completely or partially buried underground.
The peridium, the outer layer of the egg, is white, or purple/red, with two or three layers. The outer layer is thin, membranous, and elastic, while the inner layer is thicker, gelatinous, and continuous. At maturity, the peridium opens up and remains as a volva at the base of the receptaculum.
The fertile portion of the fruiting body is often borne on the end of a wide, fleshy or spongy stalk (as in the Phallales), which may be cylindrical, star-shaped, or reticulate (forming a network).
They may be brightly colored, sometimes with a lattice or veil-like membrane enclosing and protecting the spores. The spore-containing substance, the gleba, is typically gelatinous, often foul-smelling, and deliquescent (becoming liquid from the absorption of water). The gleba is formed on the exterior face of the cap or the upper part of the receptacle.
The basidia are small and narrowly club-shaped or fusiform, short-lived (evanescent), with four to eight sterigmata. The spores are usually ellipsoid or cylindrical in shape, hyaline or pale brown, smooth, more or less smooth-walled, and truncated at the base.
Phallus indusiatus, commonly called the bamboo mushrooms, bamboo pith, long net stinkhorn, crinoline stinkhorn or veiled lady, is a fungus in the family Phallaceae, or stinkhorns.
It has a cosmopolitan distribution in tropical areas, and is found in southern Asia, Africa, the Americas, and Australia, where it grows in woodlands and gardens in rich soil and well-rotted woody material.
The fruit body of the fungus is characterised by a conical to bell-shaped cap on a stalk and a delicate lacy "skirt", or indusium, that hangs from beneath the cap and reaches nearly to the ground. First described scientifically in 1798 by French botanist Étienne Pierre Ventenat, the species has often been referred to a separate genus Dictyophora along with other Phallus
\\\\nspecies featuring an indusium.
P. indusiatus can be distinguished from other similar species by differences in distribution, size, color, and indusium length. Mature fruit bodies are up to 25 cm (10 in) tall with a conical to bell-shaped cap that is 1.5–4 cm (0.6–1.6 in) wide. The cap is covered with a greenish-brown spore-containing slime, which attracts flies and other insects that eat the spores and disperse them.
An edible mushroom featured as an ingredient in Chinese haute cuisine, it is used in stir-fries and chicken soups. The mushroom, grown commercially and commonly sold in Asian markets, is rich in protein, carbohydrates, and dietary fiber. The mushroom also contains various bioactive compounds, and has antioxidant and antimicrobial properties.
Phallus indusiatus has a recorded history of use in Chinese medicine extending back to the 7th century AD, and features in Nigerian folklore.
Phallus indusiatus has many common names based on its appearance, including long net stinkhorn, crinoline stinkhorn, basket stinkhorn, bridal veil fungus, and veiled lady. A Chinese common name that alludes to its typical growth habitat is "bamboo mushroom" (simplified Chinese: 竹荪; traditional Chinese: 竹蓀; pinyin: zhúsūn).
Immature fruit bodies of P. indusiatus are initially enclosed in an egg-shaped to roughly spherical subterranean structure encased in a peridium.
The "egg" ranges in color from whitish to buff to reddish-brown, measures up to 6 cm (2.4 in) in diameter, and usually has a thick mycelial cord attached at the bottom. As the mushroom matures, the pressure caused by the enlargement of the internal structures cause the peridium to tear and the fruit body rapidly emerges from the "egg". The mature mushroom is up to 25 cm (9.8 in) tall and girded with a net-like structure called the indusium (or less technically a "skirt") that hangs down from the conical to bell-shaped cap.
The netlike openings of the indusium may be polygonal or round in shape. Well-developed specimens have an indusium that reaches to the volva and flares out somewhat before collapsing on the stalk.
The cap is 1.5–4 cm (0.6–1.6 in) wide and its reticulated (pitted and ridged) surface is covered with a layer of greenish-brown and foul-smelling slime, the gleba, which initially partially obscures the reticulations.
The top of the cap has a small hole. The stalk is 7–25 cm (2.8–9.8 in) long and 1.5–3 cm (0.6–1.2 in) thick. The hollow stalk is white, roughly equal in width throughout its length, sometimes curved, and spongy.
The ruptured peridium remains as a loose volva at the base of the stalk. Fruit bodies develop during the night and require 10–15 hours to fully develop after emerging from the peridium. They are short-lived, typically lasting no more than a few days. At that point the slime has usually been removed by insects, leaving the pale off-white, bare cap surface exposed.
Spores of P. indusiatus are thin-walled, smooth, elliptical or slightly curved, hyaline (translucent), and measure 2–3 by 1–1.5 μm.
Phallus multicolor is similar in overall appearance, but it has a more brightly coloured cap, stem and indusium, and it is usually smaller. It is found in Australia, Guam, Sumatra, Java, Borneo, Papua New Guinea, Zaire, and Tobago as well as Hawaii.
In contrast, the cap surface of P. indusiatus tends to have conspicuous reticulations that remain clearly visible under the gleba.
P. indusiatus is saprobic deriving nutrients from breaking down wood and plant organic matter. The fruit bodies grow singly or in groups in disturbed ground and among wood chips.
In eastern Asia, P. indusiatus is considered a delicacy and an aphrodisiac. Previously only collected in the wild, where it is not abundant, it was difficult to procure.
Phallus indusiatus has been cultivated on a commercial scale in China since 1979. In the Fujian Province of China known for a thriving mushroom industry that cultivates 45 species of edible fungi, P. indusiatus is produced in the counties of Fuan, Jianou, and Ningde.
The optimal temperature for the growth of mushroom spawn and fruit bodies is about 24 °C (75 °F), with a relative humidity of 90–95%. Other substrates that can be used for the cultivation of the fungus include bamboo leaves and small stems, soybean pods or stems, corn stems, and willow leaves.
Nutritional analyses of P. indusiatus show that the fruit bodies are over 90% water, about 6% fiber, 4.8% protein, 4.7% fat, and several mineral elements, including calcium, although the mineral composition in the fungus may depend on corresponding concentrations in the growth substrate.
Medicinal properties have been ascribed to Phallus indusiatus from the time of the Chinese Tang Dynasty when it was described in pharmacopoeia.
The fungus was used to treat many inflammatory,
stomach, and neural diseases. Southern China's Miao people continue to use it traditionally for a number of afflictions, including injuries and pains, cough,
dysentery, enteritis, leukemia, and feebleness, and it has been prescribed clinically as a treatment for laryngitis, leucorrhea, fever, and oliguria (low urine output), diarrhea, hypertension, cough, hyperlipidemia, and in anticancer therapy. Modern science has probed the biochemical basis of these putative medicinal benefits.
The fungus has long been recognised to have
antibacterial properties: the addition of the fungus to soup broth was known to prevent it from spoiling for several days.
One of the responsible antibiotics, albaflavenone, was isolated in 2011. It is a sesquiterpenoid that was already known from the soil bacterium Streptomyces albidoflavus. Experiments have shown that extracts of P. indusiatus have antioxidant in addition to antimicrobial properties.
Kingdom : Fungi
Division : Basidiomycota
Class :vAgaricomycetes
Order : Phallales
Family : Phallaceae
Genus : Phallus
Species : P. indusiatus
Binomial name Phallus indusiatus
Vent. (1798)
Synonyms :
Dictyophora indusiata (Vent.) Desv. (1809)
Hymenophallus indusiatus (Vent.) Nees (1817)




Cantharellus lateritius
smooth chanterelle / edible
smooth chanterelle / edible

Cantharellus lateritius, commonly known as the smooth chanterelle, is a species of edible fungus in the mushroom family Cantharellaceae.
An ectomycorrhizal species, it is found in Asia, Africa, and North America. The species has a complex taxonomic history, and has undergone several name changes since its first description by American mycologist Lewis David de Schweinitz in 1822.
The fruit bodies of the fungus are brightly colored yellow to orange, and usually highly conspicuous against the soil in which they are found.
At maturity, the mushroom resembles a filled funnel with the spore-bearing surface along the sloping outer sides. The texture of the fertile undersurface (hymenium) of the caps is a distinguishing characteristic of the species: unlike the well-known golden chanterelle, the hymenium of C. lateritius is much smoother.
Chemical analysis has revealed the presence of several carotenoid compounds in the fruit bodies.
The caps of the C. lateritius fruiting bodies typically range between 2 to 9 cm (0.8 to 3.5 in) in diameter, with a flattened to somewhat funnel-shaped top surface and a wavy margin, cap surface is dry, slightly tomentose (covered with a layer of fine hairs), and a deep and bright orange-yellow color, with older specimens fading to more yellow in age.
Distinctive margins of the cap are a paler yellow, and typically curve downward in young specimens. Fruiting bodies can reach a height of 12 cm (4.7 in).
The hymenophore (the spore-bearing surface) is initially smooth and without wrinkles, but gradually develops channels or ridges, and what appear to be very shallow gills that are vein-like, and less than 1 mm wide. The color is pale yellow, and is continuous with the surface of the stem.
The stem is rather plump and stout, 1.5 to 4.5 cm (0.6 to 1.8 in) long and 0.5 to 1.7 cm (0.2 to 0.7 in) thick, more or less cylindrical, tapering downwards towards the base. Internally, the stems are either stuffed (filled with cotton-like mycelia) or solid.
Rarely, fruiting bodies may be clumped together with stems conjoined at the base; in these cases there are usually no more than three fused stems.
The flesh is solid to partly hollow (sometimes due to insect larvae), with a pale yellow color, it is 0.5 to 0.9 cm (0.2 to 0.4 in) thick.
The spores are smooth, with a roughly ellipsoid shape, and have typical dimensions of 7–7.5 by 4.5–5 µm.
In deposit, such as in a spore print, the spores are light yellow orange, while under the microscope they are a very pale yellowish.
The spore bearing cells—the basidia—are 75–80 by 7–9 µm, 4-5-6-spored, slightly club-shaped, and with a distinctly thickened wall at the base.
Clamp connections (short branches connecting one cell to the previous cell to allow passage of the products of nuclear division) are present in the hyphae of all parts of the fruiting body.
Cantharellus lateritius is pinker than the golden chanterelle (C. cibarius) and has thicker flesh in addition to the smoother hymenial surface.
C. odoratus is also similar in appearance, and is distinguished by a thinner flesh and a hollow stem.
The poisonous "Jack O'Lantern" mushroom, Omphalotus olearius, is roughly similar in stature and color, but can be differentiated from C. lateritius by its true gills with sharply defined edges, and growth on decaying wood (although the wood may be buried in the soil) usually in large overlapping clusters.
One author considers Cantharellus lateritius to likely represent a species complex, including "all the chanterelles with a completely smooth hymenophore, sweet smell, and clamped hyphae."
Like all species in the genus Cantharellus, C. lateritius is edible, and often considered choice. The odor resembles apricots, and the taste is mild or "moderately to faintly acrid".
In the opinion of McFarland and Mueller, authors of a field guide to edible fungi of Illinois, compared to the well-known C. cibarius, C. lateritius is "in general somewhat disappointing when compared with their delicious relatives".
Cantharellus lateritius is distributed in North America, Africa, Malaysia and the Himalayas (specifically, the Almora hills in Uttar Pradesh). In the United States, its range extends northward to Michigan and New England.
Typically found growing solitary, in groups or in clusters under hardwood trees, the fungus produces fruit bodies in the summer and autumn.
In the New England area of the United States, mycologist Howard Bigelow has noted it to grow on road shoulders in grass near oaks, it also has a predilection for growing on sloping creek banks.
In Malaysia, it is found growing on the soil in forests, mostly under species of Shorea (rainforest trees in the family Dipterocarpaceae).
C. lateritius has been reported from the Western Ghats, Kerala, India, forming ectomycorrhizal association with endemic tree species like Vateria indica, Hopea parviflora, Diospyros malabarica, Myristica malabarica in semi-evergreen to evergreen forests.
Kingdom: Fungi
Division: Basidiomycota
Class: Agaricomycetes
Order: Cantharellales
Family: Cantharellaceae
Genus: Cantharellus
Species: C. lateritius
Binomial name Cantharellus lateritius
(Berk.) Singer (1951)
An ectomycorrhizal species, it is found in Asia, Africa, and North America. The species has a complex taxonomic history, and has undergone several name changes since its first description by American mycologist Lewis David de Schweinitz in 1822.
The fruit bodies of the fungus are brightly colored yellow to orange, and usually highly conspicuous against the soil in which they are found.
At maturity, the mushroom resembles a filled funnel with the spore-bearing surface along the sloping outer sides. The texture of the fertile undersurface (hymenium) of the caps is a distinguishing characteristic of the species: unlike the well-known golden chanterelle, the hymenium of C. lateritius is much smoother.
Chemical analysis has revealed the presence of several carotenoid compounds in the fruit bodies.
The caps of the C. lateritius fruiting bodies typically range between 2 to 9 cm (0.8 to 3.5 in) in diameter, with a flattened to somewhat funnel-shaped top surface and a wavy margin, cap surface is dry, slightly tomentose (covered with a layer of fine hairs), and a deep and bright orange-yellow color, with older specimens fading to more yellow in age.
Distinctive margins of the cap are a paler yellow, and typically curve downward in young specimens. Fruiting bodies can reach a height of 12 cm (4.7 in).
The hymenophore (the spore-bearing surface) is initially smooth and without wrinkles, but gradually develops channels or ridges, and what appear to be very shallow gills that are vein-like, and less than 1 mm wide. The color is pale yellow, and is continuous with the surface of the stem.
The stem is rather plump and stout, 1.5 to 4.5 cm (0.6 to 1.8 in) long and 0.5 to 1.7 cm (0.2 to 0.7 in) thick, more or less cylindrical, tapering downwards towards the base. Internally, the stems are either stuffed (filled with cotton-like mycelia) or solid.
Rarely, fruiting bodies may be clumped together with stems conjoined at the base; in these cases there are usually no more than three fused stems.
The flesh is solid to partly hollow (sometimes due to insect larvae), with a pale yellow color, it is 0.5 to 0.9 cm (0.2 to 0.4 in) thick.
The spores are smooth, with a roughly ellipsoid shape, and have typical dimensions of 7–7.5 by 4.5–5 µm.
In deposit, such as in a spore print, the spores are light yellow orange, while under the microscope they are a very pale yellowish.
The spore bearing cells—the basidia—are 75–80 by 7–9 µm, 4-5-6-spored, slightly club-shaped, and with a distinctly thickened wall at the base.
Clamp connections (short branches connecting one cell to the previous cell to allow passage of the products of nuclear division) are present in the hyphae of all parts of the fruiting body.
Cantharellus lateritius is pinker than the golden chanterelle (C. cibarius) and has thicker flesh in addition to the smoother hymenial surface.
C. odoratus is also similar in appearance, and is distinguished by a thinner flesh and a hollow stem.
The poisonous "Jack O'Lantern" mushroom, Omphalotus olearius, is roughly similar in stature and color, but can be differentiated from C. lateritius by its true gills with sharply defined edges, and growth on decaying wood (although the wood may be buried in the soil) usually in large overlapping clusters.
One author considers Cantharellus lateritius to likely represent a species complex, including "all the chanterelles with a completely smooth hymenophore, sweet smell, and clamped hyphae."
Like all species in the genus Cantharellus, C. lateritius is edible, and often considered choice. The odor resembles apricots, and the taste is mild or "moderately to faintly acrid".
In the opinion of McFarland and Mueller, authors of a field guide to edible fungi of Illinois, compared to the well-known C. cibarius, C. lateritius is "in general somewhat disappointing when compared with their delicious relatives".
Cantharellus lateritius is distributed in North America, Africa, Malaysia and the Himalayas (specifically, the Almora hills in Uttar Pradesh). In the United States, its range extends northward to Michigan and New England.
Typically found growing solitary, in groups or in clusters under hardwood trees, the fungus produces fruit bodies in the summer and autumn.
In the New England area of the United States, mycologist Howard Bigelow has noted it to grow on road shoulders in grass near oaks, it also has a predilection for growing on sloping creek banks.
In Malaysia, it is found growing on the soil in forests, mostly under species of Shorea (rainforest trees in the family Dipterocarpaceae).
C. lateritius has been reported from the Western Ghats, Kerala, India, forming ectomycorrhizal association with endemic tree species like Vateria indica, Hopea parviflora, Diospyros malabarica, Myristica malabarica in semi-evergreen to evergreen forests.
Kingdom: Fungi
Division: Basidiomycota
Class: Agaricomycetes
Order: Cantharellales
Family: Cantharellaceae
Genus: Cantharellus
Species: C. lateritius
Binomial name Cantharellus lateritius
(Berk.) Singer (1951)




Clavulinopsis laeticolor
coral mushroom
coral mushroom

Clavulinopsis is a genus of coral fungi in the family Clavariaceae. The genus, first described scientifically by Casper van Overeem in 1923, has a widespread distribution.
Similar species include Clavulinopsis fusiformis,
C. helvola, Alloclavaria purpurea, Calocera cornea, Clavaria fragilis, and Macrotyphula juncea. Some cannot be distinguished without observation of microscopic features.
Other names: Handsome Club, Golden Fairy-Club.
Clavulinopsis laeticolor is a coral mushroom in the family Clavariaceae. It has fruit bodies with slender, bright orange to yellow arms up to 5 cm (2 in) tall and 3 mm wide. It fruits singly or in loose groups on the ground, often among mosses. A widely distributed species, it is found in Asia, Europe, North America, and New Zealand.
Clavulinopsis laeticolor Mushroom Identification Ecology Presumably saprobic; growing alone, scattered, gregariously, or in loose groups under hardwoods or conifers; usually terrestrial but occasionally appearing on well-rotted, moss-covered stumps; summer and fall (also winter in warmer climates); originally described from Cuba; widely distributed in North America; also documented from Central America, South America, Europe, Asia, and Oceania.
Fruiting Body 17–50 mm high; 1–4 mm wide; cylindrical and unbranched; sometimes somewhat flattened, or with a groove or a twist; dry; bald; bright orange or yellow; fading with age; whitish at the extreme base; at maturity often with a somewhat pointed tip that ages or discolors somewhat reddish or orange.
Microscopic Features: Spores 5–75–7 x 3.5–5 µ; irregularly subellipsoid to subamygdaliform, with a large protruding apiculus; smooth; hyaline and often uniguttulate in KOH; inamyloid. Basidia 35–55 x 5–8 µm; subclavate; 4-sterigmate. Cystidia not found. Contextual hyphae 3–5 µm wide; smooth; thin-walled; hyaline to golden in KOH; with small clamp connections.
Kingdom: Fungi
Division: Basidiomycota
Class: Agaricomycetes
Order: Agaricales
Family: Clavariaceae
Genus: Clavulinopsis
Species: C. laeticolor
Binomial name Clavulinopsis laeticolor
(Berk. & M.A.Curtis) R.H.Petersen (1965)
Similar species include Clavulinopsis fusiformis,
C. helvola, Alloclavaria purpurea, Calocera cornea, Clavaria fragilis, and Macrotyphula juncea. Some cannot be distinguished without observation of microscopic features.
Other names: Handsome Club, Golden Fairy-Club.
Clavulinopsis laeticolor is a coral mushroom in the family Clavariaceae. It has fruit bodies with slender, bright orange to yellow arms up to 5 cm (2 in) tall and 3 mm wide. It fruits singly or in loose groups on the ground, often among mosses. A widely distributed species, it is found in Asia, Europe, North America, and New Zealand.
Clavulinopsis laeticolor Mushroom Identification Ecology Presumably saprobic; growing alone, scattered, gregariously, or in loose groups under hardwoods or conifers; usually terrestrial but occasionally appearing on well-rotted, moss-covered stumps; summer and fall (also winter in warmer climates); originally described from Cuba; widely distributed in North America; also documented from Central America, South America, Europe, Asia, and Oceania.
Fruiting Body 17–50 mm high; 1–4 mm wide; cylindrical and unbranched; sometimes somewhat flattened, or with a groove or a twist; dry; bald; bright orange or yellow; fading with age; whitish at the extreme base; at maturity often with a somewhat pointed tip that ages or discolors somewhat reddish or orange.
Microscopic Features: Spores 5–75–7 x 3.5–5 µ; irregularly subellipsoid to subamygdaliform, with a large protruding apiculus; smooth; hyaline and often uniguttulate in KOH; inamyloid. Basidia 35–55 x 5–8 µm; subclavate; 4-sterigmate. Cystidia not found. Contextual hyphae 3–5 µm wide; smooth; thin-walled; hyaline to golden in KOH; with small clamp connections.
Kingdom: Fungi
Division: Basidiomycota
Class: Agaricomycetes
Order: Agaricales
Family: Clavariaceae
Genus: Clavulinopsis
Species: C. laeticolor
Binomial name Clavulinopsis laeticolor
(Berk. & M.A.Curtis) R.H.Petersen (1965)


Alloclavaria purpurea

Alloclavaria is a clavarioid genus in the Hymenochaetales recently segregated from Clavaria by molecular analysis.
Phylogenetically related fungi are in the agaricoid genera Rickenella, Contumyces, Gyroflexus, Loreleia,
Cantharellopsis and Blasiphalia, as well as the stipitate stereoid genera Cotylidia and Muscinupta.
The only species as yet placed in Alloclavaria is the type, formerly known as Clavaria purpurea under which name it is often cited or illustrated, It is suspected, via circumstantial evidence, i.e. habitat, but not proven, that Alloclavaria is mycorrhizal.
Alloclavaria purpurea is a coral fungus commonly known as the purple coral, or the purple fairy club. Formerly known as Clavaria purpurea, it has been moved to its own genus as a result of phylogenetic analysis.
The fruiting body of Alloclavaria purpurea is made of numerous slender cylindrical spindles that may grow to a height of 12 centimetres (4+3⁄4 in), with individual spindles being 2–6 millimeters thick. The color is purple or lavender, although the color fades to tan in older specimens.
The spore print is white. It is reportedly edible but
insubstantial. Fruit bodies are found in spruce-fir forests.
Division: Basidiomycota
Class: Agaricomycetes
Order: Hymenochaetales
Family: Repetobasidiaceae
Genus: Alloclavaria
Species: A. purpurea
Binomial name Alloclavaria purpurea
(Fr.) Dentinger & D.J.McLaughlin (2007)
Phylogenetically related fungi are in the agaricoid genera Rickenella, Contumyces, Gyroflexus, Loreleia,
Cantharellopsis and Blasiphalia, as well as the stipitate stereoid genera Cotylidia and Muscinupta.
The only species as yet placed in Alloclavaria is the type, formerly known as Clavaria purpurea under which name it is often cited or illustrated, It is suspected, via circumstantial evidence, i.e. habitat, but not proven, that Alloclavaria is mycorrhizal.
Alloclavaria purpurea is a coral fungus commonly known as the purple coral, or the purple fairy club. Formerly known as Clavaria purpurea, it has been moved to its own genus as a result of phylogenetic analysis.
The fruiting body of Alloclavaria purpurea is made of numerous slender cylindrical spindles that may grow to a height of 12 centimetres (4+3⁄4 in), with individual spindles being 2–6 millimeters thick. The color is purple or lavender, although the color fades to tan in older specimens.
The spore print is white. It is reportedly edible but
insubstantial. Fruit bodies are found in spruce-fir forests.
Division: Basidiomycota
Class: Agaricomycetes
Order: Hymenochaetales
Family: Repetobasidiaceae
Genus: Alloclavaria
Species: A. purpurea
Binomial name Alloclavaria purpurea
(Fr.) Dentinger & D.J.McLaughlin (2007)

Xylaria polymorpha
dead man's fingers
dead man's fingers

Xylaria is a genus of ascomycetous fungi commonly found growing on dead wood. The name comes from the Greek xýlon meaning wood.
Xylaria polymorpha, commonly known as dead man's fingers, is a saprobic fungus.
It is a common inhabitant of forest and woodland areas, usually growing from the bases of rotting or injured tree stumps and decaying wood. It has also been known to colonize substrates like woody legume pods, petioles, and herbaceous stems.
It is characterized by its elongated upright, clavate, or strap-like stromata poking up through the ground, much like fingers.
The genus Xylaria contains about 100 species of cosmopolitan fungi. Polymorpha means "many forms". As its name suggests, it has a very variable but often club-shaped fruiting body (stroma) resembling burned wood.
Often this fungus is found with a multitude of separate "digits" but at times the individual parts will be fused together.
Belonging to the phylum of fungus known as Ascomycetes (division Mycota) known as the sac fungi, they are characterized by a saclike structure, the ascus, which contains anything from four to eight ascospores in the sexual stage.
The sac fungi are separated into subgroups based on whether asci arise singly or are borne in one of several types of fruiting structures, or ascocarps, and on the method of discharge of the ascospores.
Many ascomycetes are plant pathogens, some are animal pathogens, a few are edible mushrooms, and many live on dead organic matter (as saprobes).
The largest and most commonly known ascomycetes include the morel and the truffle, however, X. polymorpha is an inedible variety.
The dark fruiting body (often black or brown, but sometimes shades of blue/green) is white on the inside, with a blackened dotted area all around.
This blackened surrounding area is made up of tiny structures called perithecia. The perithecia hold a layer of asci which contain the ascospores. The asci elongate into the ostiole, and discharge the ascospores outward.
The spore distribution is a lengthy process, sometimes taking several months to complete this part of the life cycle, this is not a common trait amongst fungi, as is normally a much swifter process.
In springtime this fungus often produces a layer of white or bluish asexual spores called conidia, which grow on its surface and surrounding area.
Division: Ascomycota
Class: Sordariomycetes
Order: Xylariales
Family: Xylariaceae
Genus: Xylaria
Species: X. polymorpha
Binomial name Xylaria polymorpha
(Pers.) Grev., (1824)
Xylaria polymorpha, commonly known as dead man's fingers, is a saprobic fungus.
It is a common inhabitant of forest and woodland areas, usually growing from the bases of rotting or injured tree stumps and decaying wood. It has also been known to colonize substrates like woody legume pods, petioles, and herbaceous stems.
It is characterized by its elongated upright, clavate, or strap-like stromata poking up through the ground, much like fingers.
The genus Xylaria contains about 100 species of cosmopolitan fungi. Polymorpha means "many forms". As its name suggests, it has a very variable but often club-shaped fruiting body (stroma) resembling burned wood.
Often this fungus is found with a multitude of separate "digits" but at times the individual parts will be fused together.
Belonging to the phylum of fungus known as Ascomycetes (division Mycota) known as the sac fungi, they are characterized by a saclike structure, the ascus, which contains anything from four to eight ascospores in the sexual stage.
The sac fungi are separated into subgroups based on whether asci arise singly or are borne in one of several types of fruiting structures, or ascocarps, and on the method of discharge of the ascospores.
Many ascomycetes are plant pathogens, some are animal pathogens, a few are edible mushrooms, and many live on dead organic matter (as saprobes).
The largest and most commonly known ascomycetes include the morel and the truffle, however, X. polymorpha is an inedible variety.
The dark fruiting body (often black or brown, but sometimes shades of blue/green) is white on the inside, with a blackened dotted area all around.
This blackened surrounding area is made up of tiny structures called perithecia. The perithecia hold a layer of asci which contain the ascospores. The asci elongate into the ostiole, and discharge the ascospores outward.
The spore distribution is a lengthy process, sometimes taking several months to complete this part of the life cycle, this is not a common trait amongst fungi, as is normally a much swifter process.
In springtime this fungus often produces a layer of white or bluish asexual spores called conidia, which grow on its surface and surrounding area.
Division: Ascomycota
Class: Sordariomycetes
Order: Xylariales
Family: Xylariaceae
Genus: Xylaria
Species: X. polymorpha
Binomial name Xylaria polymorpha
(Pers.) Grev., (1824)


Amauroderma rugosum
Black Lingzhi Mushroom
Black Lingzhi Mushroom

Amauroderma is a genus of polypore fungi in the family Ganodermataceae. The genus, widespread in tropical areas, contains about 70 species.
Amauroderma fungi are wood-decay fungi that feed and fruit on decayed branches and trunks.
The fruit bodies of Amauroderma fungi comprise a cap and a stipe, and are typically woody, leathery, or corky in texture.
The spores produced are usually spherical or nearly so, with a characteristic double wall structure that features U-shaped thickenings.
The fruit bodies of Amauroderma species are stipitate except in A. andina and may attain various shapes although centrally stipitate basidiocarps are most common. Several stipes may arise from the same base, frequently resulting in fused caps and compound fruit bodies.
In section some fruit bodies are distinct with one or two distinct inner black bands or zones. The stipe is often duplex with an outer dense layer surrounding an inner softer or hollow core sometimes separated by a black band.
In species with a distinct tomentum on the stipe, there is often a dark zone just below the tomentum of the cap. These zones are absent from some species with a pale stipe without a tomentum. However, when present they continue into the context and frequently there is also another zone stretching more or less horizontally across the context.
Most basidiospores of Amauroderma mushrooms have an inner ornamented wall on which there is a hyaline (translucent) epicutis, which is very thin and difficult to see in ordinary microscopic preparations.
Mature basidiospores are pale-yellowish. An apiculus (a depressed area where the spore was once attached to the basidium via the sterigma) is often difficult to observe.
Amauroderma rugosum (Blume & T. Nees) Torrend (Ganodermataceae) is an edible mushroom with medicinal properties. However, the effects of A. rugosum on gastric ulcer remain unclear.
Amauroderma rugosum is a normally widespread in other regions but has been evaluated as a least concern (LC). This wild medicinal fungus always be consumed by indigenous communities in some countries.
A Mycochemical Investigation of the Black Lingzhi Mushroom, Amauroderma rugosum (Agaricomycetes), Reveals Several Lipidic Compounds with Anti-Inflammatory and Antiproliferative Activities.
(Shao-Dan Chen et al. Int J Med Mushrooms. 2021)
Amauroderma rugosum commonly known as "Jiǎzī" in China is one of the traditional Chinese medicinal mushrooms and is used to reduce inflammation, treat diuretic and upset stomach, and prevent cancer.
Collecting of this wild medicine fungus by human (Indigenous communities) and deforestation of diterocarp trees widely will reduce the population of fungus and degrade their natural habitat particulary in tropical rainforest regions in Thailand, Malaysia, Singapore, Indonesia and Papua New Guinea.
Population of species worldwide are facing habitat homogenization due to human activities especially in Asia. A low population density for this species in most regions due to extremely unfavorable habitats (substrate unfavourable for this saprophyte fungus) and habitat quality reduction then the species shows declines in some countries. Unclear of current trends of this species can be observed in most regions.
The species is growing as saprophytic (wood-decay fungi) on the ground with other plants and common in lowland dipterocarp of tropic forest and sub-tropic forest.
The most suitable elevation for the growth and development of Amauroderma rugosum within on elevation of 200-1400m a.s.l. The higher humidity than 90% is the best for the growth and development of this species (Nguyen & Khanh, 2017).
Amauroderma rugosum is a wild medicinal mushroom commonly sold in China and Malaysia which has antioxidant and anti-tumor activities.
It is also worn as a necklace by the indigenous communities in Malaysia to prevent epileptic episodes and incessant crying by babies.
Division: Basidiomycota
Class: Agaricomycetes
Order: Polyporales
Family: Ganodermataceae
Genus: Amauroderma
Murrill (1905)
Species : Amauroderma rugosum
(Blume & T. Nees) Torrend, 1920
Amauroderma fungi are wood-decay fungi that feed and fruit on decayed branches and trunks.
The fruit bodies of Amauroderma fungi comprise a cap and a stipe, and are typically woody, leathery, or corky in texture.
The spores produced are usually spherical or nearly so, with a characteristic double wall structure that features U-shaped thickenings.
The fruit bodies of Amauroderma species are stipitate except in A. andina and may attain various shapes although centrally stipitate basidiocarps are most common. Several stipes may arise from the same base, frequently resulting in fused caps and compound fruit bodies.
In section some fruit bodies are distinct with one or two distinct inner black bands or zones. The stipe is often duplex with an outer dense layer surrounding an inner softer or hollow core sometimes separated by a black band.
In species with a distinct tomentum on the stipe, there is often a dark zone just below the tomentum of the cap. These zones are absent from some species with a pale stipe without a tomentum. However, when present they continue into the context and frequently there is also another zone stretching more or less horizontally across the context.
Most basidiospores of Amauroderma mushrooms have an inner ornamented wall on which there is a hyaline (translucent) epicutis, which is very thin and difficult to see in ordinary microscopic preparations.
Mature basidiospores are pale-yellowish. An apiculus (a depressed area where the spore was once attached to the basidium via the sterigma) is often difficult to observe.
Amauroderma rugosum (Blume & T. Nees) Torrend (Ganodermataceae) is an edible mushroom with medicinal properties. However, the effects of A. rugosum on gastric ulcer remain unclear.
Amauroderma rugosum is a normally widespread in other regions but has been evaluated as a least concern (LC). This wild medicinal fungus always be consumed by indigenous communities in some countries.
A Mycochemical Investigation of the Black Lingzhi Mushroom, Amauroderma rugosum (Agaricomycetes), Reveals Several Lipidic Compounds with Anti-Inflammatory and Antiproliferative Activities.
(Shao-Dan Chen et al. Int J Med Mushrooms. 2021)
Amauroderma rugosum commonly known as "Jiǎzī" in China is one of the traditional Chinese medicinal mushrooms and is used to reduce inflammation, treat diuretic and upset stomach, and prevent cancer.
Collecting of this wild medicine fungus by human (Indigenous communities) and deforestation of diterocarp trees widely will reduce the population of fungus and degrade their natural habitat particulary in tropical rainforest regions in Thailand, Malaysia, Singapore, Indonesia and Papua New Guinea.
Population of species worldwide are facing habitat homogenization due to human activities especially in Asia. A low population density for this species in most regions due to extremely unfavorable habitats (substrate unfavourable for this saprophyte fungus) and habitat quality reduction then the species shows declines in some countries. Unclear of current trends of this species can be observed in most regions.
The species is growing as saprophytic (wood-decay fungi) on the ground with other plants and common in lowland dipterocarp of tropic forest and sub-tropic forest.
The most suitable elevation for the growth and development of Amauroderma rugosum within on elevation of 200-1400m a.s.l. The higher humidity than 90% is the best for the growth and development of this species (Nguyen & Khanh, 2017).
Amauroderma rugosum is a wild medicinal mushroom commonly sold in China and Malaysia which has antioxidant and anti-tumor activities.
It is also worn as a necklace by the indigenous communities in Malaysia to prevent epileptic episodes and incessant crying by babies.
Division: Basidiomycota
Class: Agaricomycetes
Order: Polyporales
Family: Ganodermataceae
Genus: Amauroderma
Murrill (1905)
Species : Amauroderma rugosum
(Blume & T. Nees) Torrend, 1920


Physarum polycephalum,
an acellular slime mold
an acellular slime mold

Physarum is a genus of mycetozoan slime molds in the family Physaraceae.
Slime mold or slime mould is an informal name given to several kinds of unrelated eukaryotic organisms
with a life cycle that includes a free-living single-celled stage and the formation of spores.
Spores are often produced in macroscopic multicellular or multinucleate fruiting bodies which may be formed through aggregation or fusion.
Slime molds were formerly classified as fungi but are no longer considered part of that kingdom. Although not forming a single monophyletic clade, they are grouped within the paraphyletic group Protista.
More than 900 species of slime mold occur globally. Their common name refers to part of some of these organisms' life cycles where they can appear as gelatinous "slime". This is mostly seen with the Myxogastria, which are the only macroscopic slime molds.
Most slime molds are smaller than a few centimetres, but some species may reach sizes up to several square metres and masses up to 20 kilograms.
They feed on microorganisms that live in any type of dead plant material. They contribute to the decomposition of dead vegetation, and feed on bacteria and fungi.
For this reason, slime molds are usually found in soil, lawns, and on the forest floor, commonly on deciduous logs. In tropical areas they are also common on inflorescences and fruits, and in aerial situations (e.g., in the canopy of trees). In urban areas, they are found on mulch or in the leaf mold in rain gutters, and also grow in air conditioners, especially when the drain is blocked.
Physarum polycephalum, an acellular slime mold or
myxomycete popularly known as "the blob", is a protist with diverse cellular forms and broad geographic distribution.
The “acellular” moniker derives from the plasmodial stage of the life cycle: the plasmodium is a bright yellow macroscopic multinucleate coenocyte shaped
in a network of interlaced tubes. This stage of the life cycle, along with its preference for damp shady habitats, likely contributed to the original mischaracterization of the organism as a fungus.
P. polycephalum is used as a model organism for research into motility, cellular differentiation, chemotaxis, cellular compatibility, and the cell cycle.
Slime mold or slime mould is an informal name given to several kinds of unrelated eukaryotic organisms
with a life cycle that includes a free-living single-celled stage and the formation of spores.
Spores are often produced in macroscopic multicellular or multinucleate fruiting bodies which may be formed through aggregation or fusion.
Slime molds were formerly classified as fungi but are no longer considered part of that kingdom. Although not forming a single monophyletic clade, they are grouped within the paraphyletic group Protista.
More than 900 species of slime mold occur globally. Their common name refers to part of some of these organisms' life cycles where they can appear as gelatinous "slime". This is mostly seen with the Myxogastria, which are the only macroscopic slime molds.
Most slime molds are smaller than a few centimetres, but some species may reach sizes up to several square metres and masses up to 20 kilograms.
They feed on microorganisms that live in any type of dead plant material. They contribute to the decomposition of dead vegetation, and feed on bacteria and fungi.
For this reason, slime molds are usually found in soil, lawns, and on the forest floor, commonly on deciduous logs. In tropical areas they are also common on inflorescences and fruits, and in aerial situations (e.g., in the canopy of trees). In urban areas, they are found on mulch or in the leaf mold in rain gutters, and also grow in air conditioners, especially when the drain is blocked.
Physarum polycephalum, an acellular slime mold or
myxomycete popularly known as "the blob", is a protist with diverse cellular forms and broad geographic distribution.
The “acellular” moniker derives from the plasmodial stage of the life cycle: the plasmodium is a bright yellow macroscopic multinucleate coenocyte shaped
in a network of interlaced tubes. This stage of the life cycle, along with its preference for damp shady habitats, likely contributed to the original mischaracterization of the organism as a fungus.
P. polycephalum is used as a model organism for research into motility, cellular differentiation, chemotaxis, cellular compatibility, and the cell cycle.
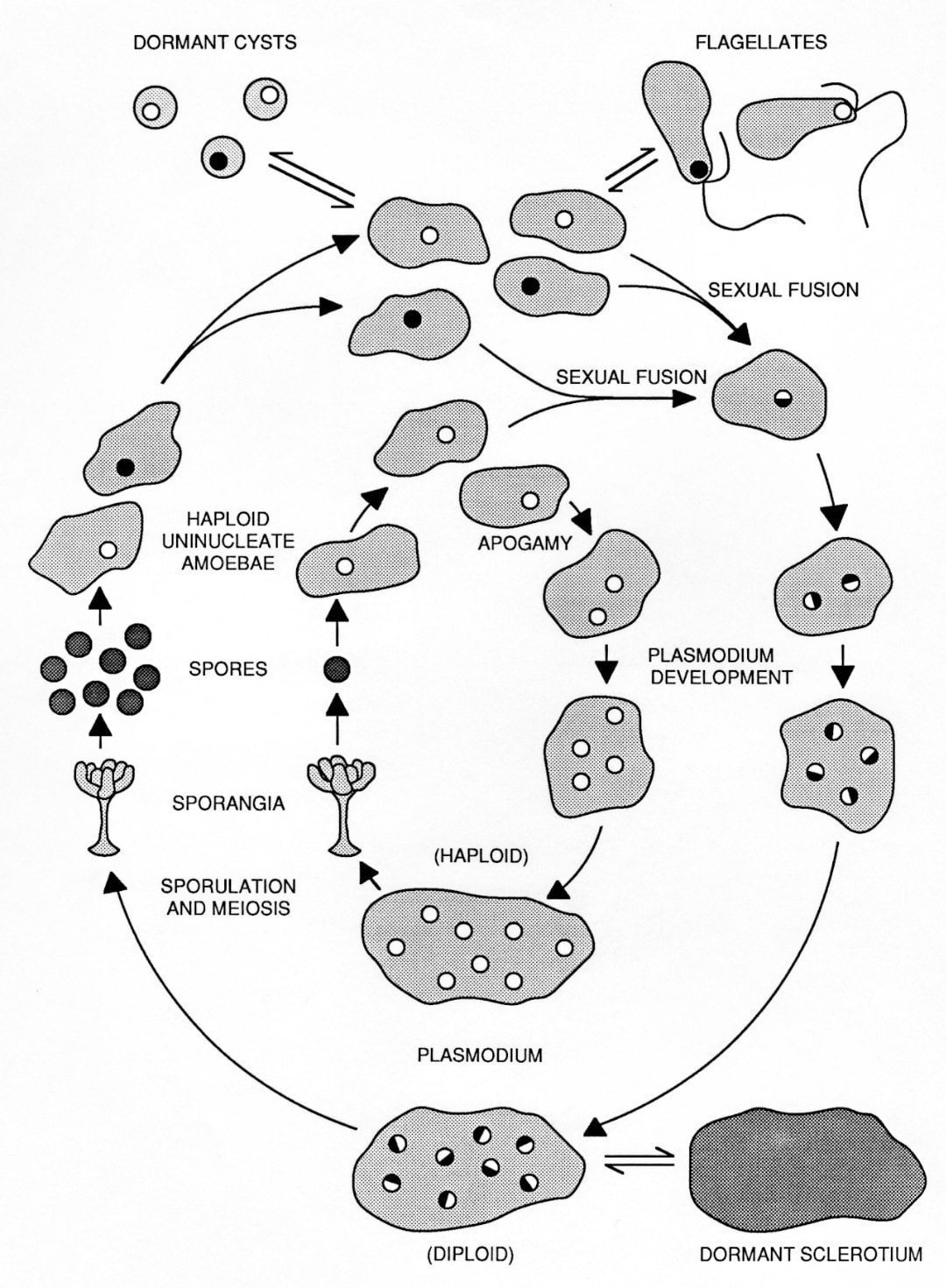
The life cycle of Physarum polycephalum. The outer circuit illustrates the natural cycle alternating between the haploid amoebal stage and diploid plasmodial stage.
The inner circuit illustrates the fully haploid "apogamic" life cycle. Both cycles exhibit all developmental stages.
The two vegetative cell types, amoebae and plasmodia, differ markedly in morphology, physiology and behavior. Amoebae are microorganisms, typically haploid, that live primarily in the soil, where they phagocytose bacteria.
In the laboratory, amoebae are grown on lawns of
live or dead Escherichia coli on nutrient agar plates, where they can multiply indefinitely.
Axenic culture of amoebae was achieved through selection of mutants capable of axenic growth. Under conditions of starvation or desiccation, the amoebae differentiate reversibly into dormant spores with cell walls.
When immersed in water, amoebae differentiate reversibly into flagellated cells, which involves a major reorganization of the cytoskeleton.
Phylum: Amoebozoa
Class: Myxogastria
Order: Physarales
Family: Physaraceae
Genus: Physarum
Species: P. polycephalum
Binomial name Physarum polycephalum
Schwein.
The inner circuit illustrates the fully haploid "apogamic" life cycle. Both cycles exhibit all developmental stages.
The two vegetative cell types, amoebae and plasmodia, differ markedly in morphology, physiology and behavior. Amoebae are microorganisms, typically haploid, that live primarily in the soil, where they phagocytose bacteria.
In the laboratory, amoebae are grown on lawns of
live or dead Escherichia coli on nutrient agar plates, where they can multiply indefinitely.
Axenic culture of amoebae was achieved through selection of mutants capable of axenic growth. Under conditions of starvation or desiccation, the amoebae differentiate reversibly into dormant spores with cell walls.
When immersed in water, amoebae differentiate reversibly into flagellated cells, which involves a major reorganization of the cytoskeleton.
Phylum: Amoebozoa
Class: Myxogastria
Order: Physarales
Family: Physaraceae
Genus: Physarum
Species: P. polycephalum
Binomial name Physarum polycephalum
Schwein.


Tremella mesenterica
witches' butter
witches' butter

Tremella mesenterica (common names include yellow brain, golden jelly fungus, yellow trembler, and witches' butter) is a common jelly fungus in the family Tremellaceae of the Agaricomycotina.
It is most frequently found on dead but attached and on recently fallen branches, especially of angiosperms
as a parasite of wood decay fungi in the genus Peniophora.
The gelatinous, orange-yellow fruit body of the fungus, which can grow up to 7.5 cm (3.0 in) diameter, has a convoluted or lobed surface that is greasy or slimy when damp. It grows in crevices in bark, appearing during rainy weather.
Within a few days after rain it dries into a thin film or shriveled mass capable of reviving after subsequent rain. This fungus occurs widely in deciduous and mixed forests and is widely distributed in temperate and tropical regions that include Africa, Asia, Australia, Europe, North and South America.
Although considered bland and flavorless, the fungus is edible. Tremella mesenterica produces carbohydrates that are attracting research interest because of their various biological activities.
The species was originally described from Sweden as Helvella mesenterica by the naturalist Jacob Christian Schäffer in 1774. Valid description was provided by Anders Jahan Retzius in 1769.
It was later (1822) sanctioned by Elias Magnus Fries in the second volume of his Systema Mycologicum. It is the type species of the genus Tremella.
Its distinctive appearance has led the species to accumulate a variety of common names, including "yellow trembler", "yellow brain", "golden jelly fungus", and "witches' butter;" although this latter name is also applied to Exidia glandulosa, its origin may stem from Swedish folklore surrounding witchcraft, in which a bile spewed up by thieving "Carriers" is referred to as, "butter of the witches."
The fruit body has an irregular shape, and usually breaks through the bark of dead branches. It is up to 7.5 cm (3.0 in) broad and 2.5 to 5.0 cm (1.0 to 2.0 in) high, rounded to variously lobed or brain-like in appearance.
The fruit body is gelatin-like but tough when wet, and hard when dry. The surface is usually smooth, the lobes translucent, deep yellow or bright yellow-orange, fading to pale yellow, rarely unpigmented and white or colorless. The fruit bodies dry to a dark reddish or orange. The spores, viewed in mass, are whitish or pale yellow.
The basidia (spore-bearing cells) are ellipsoid to roughly spherical in shape, not or rarely stalked, and typically 15–21 µm wide. They contain two to four septa that divide it into compartments; the septa are most frequently diagonal or vertical.
Asexual reproduction in T. mesenterica is carried out through the formation of spores called conidia, which arise from conidiophores—specialized hyphal cells that are morphologically distinct from the somatic hyphae.
The conidiophores are densely branched and normally abundant in the hymenium; young specimens may be entirely conidial.
The conidia are roughly spherical, ovoid, or ellipsoid, and about 2.0–3.0 by 2.0–2.5 µm. They may be so numerous that young fruit bodies may be covered in a bright yellow, conidial slime. The spores are broadly ellipsoid to oblong, on average 10.0–16.0 by 6.0–9.5 µm; they germinate by germ tube or by yeast-like conidia of identical form to the conidia produced on the conidiophores.
Although some have claimed the fungus to be inedible or merely "non-poisonous", other sources say that it is edible but flavorless.
The gelatinous to rubbery consistency lends texture to soups. In China, the fungus is used by vegetarians to prepare "an immunomodulating cooling soup with lotus seed, lily bulbs, jujube, etc."
Division: Basidiomycota
Class: Tremellomycetes
Order: Tremellales
Family: Tremellaceae
Genus: Tremella
Species: T. mesenterica
Binomial name Tremella mesenterica Retz. (1769)
It is most frequently found on dead but attached and on recently fallen branches, especially of angiosperms
as a parasite of wood decay fungi in the genus Peniophora.
The gelatinous, orange-yellow fruit body of the fungus, which can grow up to 7.5 cm (3.0 in) diameter, has a convoluted or lobed surface that is greasy or slimy when damp. It grows in crevices in bark, appearing during rainy weather.
Within a few days after rain it dries into a thin film or shriveled mass capable of reviving after subsequent rain. This fungus occurs widely in deciduous and mixed forests and is widely distributed in temperate and tropical regions that include Africa, Asia, Australia, Europe, North and South America.
Although considered bland and flavorless, the fungus is edible. Tremella mesenterica produces carbohydrates that are attracting research interest because of their various biological activities.
The species was originally described from Sweden as Helvella mesenterica by the naturalist Jacob Christian Schäffer in 1774. Valid description was provided by Anders Jahan Retzius in 1769.
It was later (1822) sanctioned by Elias Magnus Fries in the second volume of his Systema Mycologicum. It is the type species of the genus Tremella.
Its distinctive appearance has led the species to accumulate a variety of common names, including "yellow trembler", "yellow brain", "golden jelly fungus", and "witches' butter;" although this latter name is also applied to Exidia glandulosa, its origin may stem from Swedish folklore surrounding witchcraft, in which a bile spewed up by thieving "Carriers" is referred to as, "butter of the witches."
The fruit body has an irregular shape, and usually breaks through the bark of dead branches. It is up to 7.5 cm (3.0 in) broad and 2.5 to 5.0 cm (1.0 to 2.0 in) high, rounded to variously lobed or brain-like in appearance.
The fruit body is gelatin-like but tough when wet, and hard when dry. The surface is usually smooth, the lobes translucent, deep yellow or bright yellow-orange, fading to pale yellow, rarely unpigmented and white or colorless. The fruit bodies dry to a dark reddish or orange. The spores, viewed in mass, are whitish or pale yellow.
The basidia (spore-bearing cells) are ellipsoid to roughly spherical in shape, not or rarely stalked, and typically 15–21 µm wide. They contain two to four septa that divide it into compartments; the septa are most frequently diagonal or vertical.
Asexual reproduction in T. mesenterica is carried out through the formation of spores called conidia, which arise from conidiophores—specialized hyphal cells that are morphologically distinct from the somatic hyphae.
The conidiophores are densely branched and normally abundant in the hymenium; young specimens may be entirely conidial.
The conidia are roughly spherical, ovoid, or ellipsoid, and about 2.0–3.0 by 2.0–2.5 µm. They may be so numerous that young fruit bodies may be covered in a bright yellow, conidial slime. The spores are broadly ellipsoid to oblong, on average 10.0–16.0 by 6.0–9.5 µm; they germinate by germ tube or by yeast-like conidia of identical form to the conidia produced on the conidiophores.
Although some have claimed the fungus to be inedible or merely "non-poisonous", other sources say that it is edible but flavorless.
The gelatinous to rubbery consistency lends texture to soups. In China, the fungus is used by vegetarians to prepare "an immunomodulating cooling soup with lotus seed, lily bulbs, jujube, etc."
Division: Basidiomycota
Class: Tremellomycetes
Order: Tremellales
Family: Tremellaceae
Genus: Tremella
Species: T. mesenterica
Binomial name Tremella mesenterica Retz. (1769)


Podostroma cornu-damae
(Japanese: カエンタケ, Hepburn: kaentake)
poison fire coral,
(Japanese: カエンタケ, Hepburn: kaentake)
poison fire coral,


Maju forest 9 - 8 - 21
Daylight shot.
Daylight shot.
Podostroma cornu-damae (Japanese: カエンタケ, Hepburn: kaentake), also known as the poison fire coral, is a species of fungus in the family Hypocreaceae.
The fruit bodies of the fungus are highly toxic, and have been responsible for several fatalities in Japan. The fungus contains several trichothecene mycotoxins.
The species was originally described as Hypocrea cornu-damae by Narcisse Théophile Patouillard in 1895, and later transferred to the genus Podocrea in 1905 by Pier Andrea Saccardo.
In 1994 Japanese mycologists Tsuguo Hongo and Masana Izawa placed the species in the genus
Podostroma.
The fungus was once thought to be only native to Korea and Japan, but recent discoveries have been made in Java, Papua New Guinea and some parts of Australia and also found in Singapore.
The conidiophores (specialized fungal hyphae that produce conidia) are up to 400 μm high and about 2–4 μm wide in the main axial hyphae.
The phialides are arranged in tufts with narrow angles at the top, similar to the branching hyphae found in Trichoderma species. The conidia are roughly spherical with a truncate base in each spore, pale green in color, and measure 2.5–3.5 μm in diameter. Their surfaces are almost smooth, but sometimes appearing very faintly roughened with light microscopy.
Several poisonings have been reported in Japan resulting from the consumption of the fungus. In 1999, one of a group of five people from Niigata prefecture
died two days after consuming about 1 gram (0.035 oz) of fruit body that had been soaked in sake.
In 2000, an individual from Gunma prefecture died after eating the fried mushroom. Symptoms associated with consumption in these cases included stomach pains, changes in perception, decrease in the number of leukocytes and thrombocytes, peeling skin on the face, hair loss, and shrinking of the cerebellum, resulting in speech impediment and problems with voluntary movement.
In another instance, an autopsy revealed multiple organ failure, including acute kidney failure, liver necrosis and disseminated intravascular coagulation.
In one case of poisoning, the patient suffered from
hemophagocytosis, in addition to severe leukocytopenia and thrombocytopenia, seven days after ingesting the fungus. Plasmapheresis and administration of granulocyte colony-stimulating factor were used to treat the blood abnormalities.
The authors suggested that these treatments, in addition to the large volume of administered
intravenous saline - 9 liters (2.0 imp gal; 2.4 U.S. gal) over a 12-hour period—were responsible for his successful recovery.
The poisoning symptoms are similar to those observed previously with animals that had consumed
trichothecene mycotoxins. Japanese researchers detected the presence of the macrocyclic
trichothecenes satratoxin H, satratoxin H 12′,13′-diacetate, satratoxin H 12′-acetate, and satratoxin H 13′-acetate.
When grown in liquid culture the fungus additionally produces roridin E and verrucarin J. With the exception of verrucarin J, a 500-microgram dose of any of these compounds, when injected into the abdomen of mice, will result in their death the following day. It has been claimed that touching the fungus can cause skin rashes, but this is controversial.
The hymenium is the tissue layer on the hymenophore
of a fungal fruiting body where the cells develop into basidia or asci, which produce spores.
In some species all of the cells of the hymenium develop into basidia or asci, while in others some cells develop into sterile cells called cystidia
(basidiomycetes) or paraphyses (ascomycetes).
Cystidia are often important for microscopic identification. The subhymenium consists of the supportive hyphae from which the cells of the hymenium grow, beneath which is the hymenophoral
trama, the hyphae that make up the mass of the hymenophore.
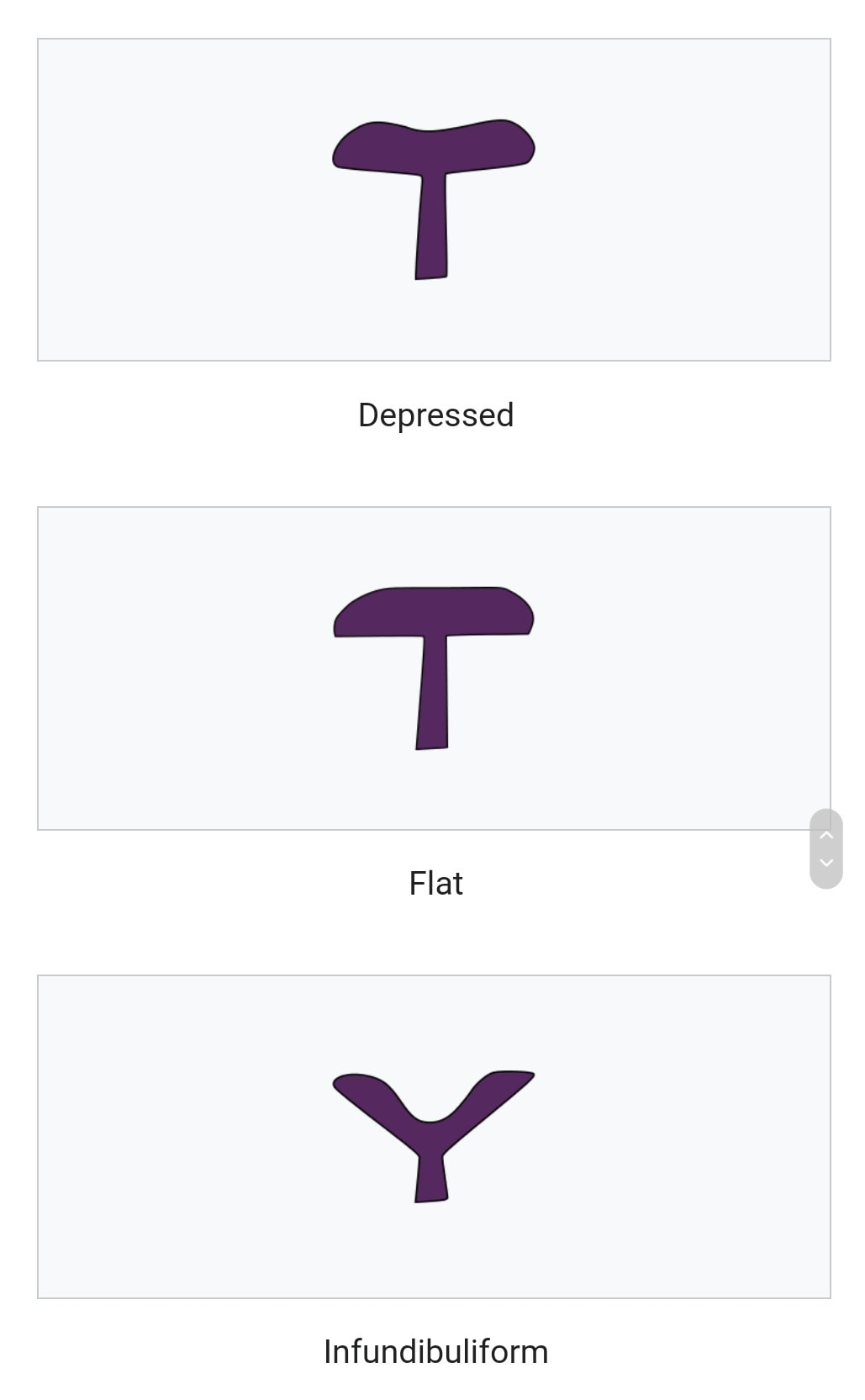
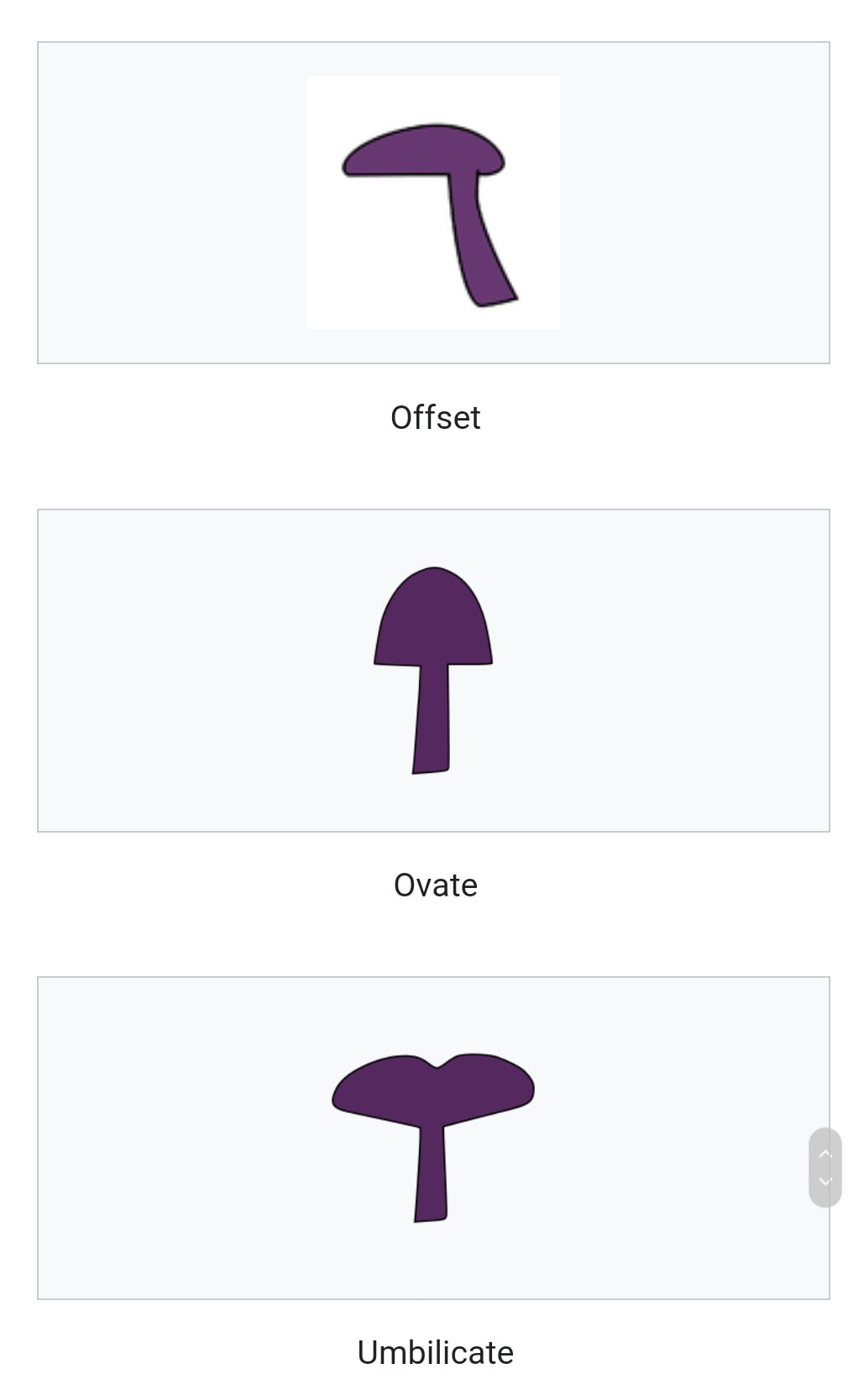
The pileus is the technical name for the cap, or cap-like part, of a basidiocarp or ascocarp (fungal fruiting body) that supports a spore-bearing surface, the hymenium.
The hymenium (hymenophore) may consist of lamellae, tubes, or teeth, on the underside of the pileus. A pileus is characteristic of agarics, boletes, some polypores, tooth fungi, and some ascomycetes.
Pilei can be formed in various shapes, and the shapes can change over the course of the developmental cycle of a fungus.
The most familiar pileus shape is hemispherical or convex. Convex pilei often continue to expand as they mature until they become flat. Many well-known species have a convex pileus, including the button mushroom, various Amanita species and boletes.
Some, such as the parasol mushroom, have distinct bosses or umbos and are described as umbonate. An umbo is a knobby protrusion at the center of the cap. Some fungi, such as chanterelles have a funnel- or trumpet-shaped appearance. In these cases the pileus is termed infundibuliform.
In mycology, a stipe is the stem or stalk-like feature supporting the cap of a mushroom. Like all tissues of the mushroom other than the hymenium, the stipe is composed of sterile hyphal tissue. In many instances, however, the fertile hymenium extends down the stipe some distance. Fungi that have stipes are said to be stipitate.
The evolutionary benefit of a stipe is generally considered to be in mediating spore dispersal. An elevated mushroom will more easily release its spores into wind currents or onto passing animals.
Nevertheless, many mushrooms do not have stipes, including cup fungi, puffballs, earthstars, some polypores, jelly fungi, ergots, and smuts.
It is often the case that features of the stipe are required to make a positive identification of a mushroom.
Such distinguishing characters include: the texture of the stipe (fibrous, brittle, chalky, leathery, firm, etc.) whether it has remains of a partial veil (such as an annulus or cortina) or universal veil (volva) whether
the stipes of many mushrooms fuse at their baseits general size and shapewhether the stipe extends underground in a root-like structure (a rhizome)
When collecting mushrooms for identification it is critical to maintain all these characters intact by digging the mushroom out of the soil, rather than cutting it off mid-stipe.
Division: Ascomycota
Class: Sordariomycetes
Order: Hypocreales
Family: Hypocreaceae
Genus: Podostroma
Species: P. cornu-damae
Binomial name Podostroma cornu-damae
(Patouillard) Hongo & Izawa
The fruit bodies of the fungus are highly toxic, and have been responsible for several fatalities in Japan. The fungus contains several trichothecene mycotoxins.
The species was originally described as Hypocrea cornu-damae by Narcisse Théophile Patouillard in 1895, and later transferred to the genus Podocrea in 1905 by Pier Andrea Saccardo.
In 1994 Japanese mycologists Tsuguo Hongo and Masana Izawa placed the species in the genus
Podostroma.
The fungus was once thought to be only native to Korea and Japan, but recent discoveries have been made in Java, Papua New Guinea and some parts of Australia and also found in Singapore.
The conidiophores (specialized fungal hyphae that produce conidia) are up to 400 μm high and about 2–4 μm wide in the main axial hyphae.
The phialides are arranged in tufts with narrow angles at the top, similar to the branching hyphae found in Trichoderma species. The conidia are roughly spherical with a truncate base in each spore, pale green in color, and measure 2.5–3.5 μm in diameter. Their surfaces are almost smooth, but sometimes appearing very faintly roughened with light microscopy.
Several poisonings have been reported in Japan resulting from the consumption of the fungus. In 1999, one of a group of five people from Niigata prefecture
died two days after consuming about 1 gram (0.035 oz) of fruit body that had been soaked in sake.
In 2000, an individual from Gunma prefecture died after eating the fried mushroom. Symptoms associated with consumption in these cases included stomach pains, changes in perception, decrease in the number of leukocytes and thrombocytes, peeling skin on the face, hair loss, and shrinking of the cerebellum, resulting in speech impediment and problems with voluntary movement.
In another instance, an autopsy revealed multiple organ failure, including acute kidney failure, liver necrosis and disseminated intravascular coagulation.
In one case of poisoning, the patient suffered from
hemophagocytosis, in addition to severe leukocytopenia and thrombocytopenia, seven days after ingesting the fungus. Plasmapheresis and administration of granulocyte colony-stimulating factor were used to treat the blood abnormalities.
The authors suggested that these treatments, in addition to the large volume of administered
intravenous saline - 9 liters (2.0 imp gal; 2.4 U.S. gal) over a 12-hour period—were responsible for his successful recovery.
The poisoning symptoms are similar to those observed previously with animals that had consumed
trichothecene mycotoxins. Japanese researchers detected the presence of the macrocyclic
trichothecenes satratoxin H, satratoxin H 12′,13′-diacetate, satratoxin H 12′-acetate, and satratoxin H 13′-acetate.
When grown in liquid culture the fungus additionally produces roridin E and verrucarin J. With the exception of verrucarin J, a 500-microgram dose of any of these compounds, when injected into the abdomen of mice, will result in their death the following day. It has been claimed that touching the fungus can cause skin rashes, but this is controversial.
The hymenium is the tissue layer on the hymenophore
of a fungal fruiting body where the cells develop into basidia or asci, which produce spores.
In some species all of the cells of the hymenium develop into basidia or asci, while in others some cells develop into sterile cells called cystidia
(basidiomycetes) or paraphyses (ascomycetes).
Cystidia are often important for microscopic identification. The subhymenium consists of the supportive hyphae from which the cells of the hymenium grow, beneath which is the hymenophoral
trama, the hyphae that make up the mass of the hymenophore.
The pileus is the technical name for the cap, or cap-like part, of a basidiocarp or ascocarp (fungal fruiting body) that supports a spore-bearing surface, the hymenium.
The hymenium (hymenophore) may consist of lamellae, tubes, or teeth, on the underside of the pileus. A pileus is characteristic of agarics, boletes, some polypores, tooth fungi, and some ascomycetes.
Pilei can be formed in various shapes, and the shapes can change over the course of the developmental cycle of a fungus.
The most familiar pileus shape is hemispherical or convex. Convex pilei often continue to expand as they mature until they become flat. Many well-known species have a convex pileus, including the button mushroom, various Amanita species and boletes.
Some, such as the parasol mushroom, have distinct bosses or umbos and are described as umbonate. An umbo is a knobby protrusion at the center of the cap. Some fungi, such as chanterelles have a funnel- or trumpet-shaped appearance. In these cases the pileus is termed infundibuliform.
In mycology, a stipe is the stem or stalk-like feature supporting the cap of a mushroom. Like all tissues of the mushroom other than the hymenium, the stipe is composed of sterile hyphal tissue. In many instances, however, the fertile hymenium extends down the stipe some distance. Fungi that have stipes are said to be stipitate.
The evolutionary benefit of a stipe is generally considered to be in mediating spore dispersal. An elevated mushroom will more easily release its spores into wind currents or onto passing animals.
Nevertheless, many mushrooms do not have stipes, including cup fungi, puffballs, earthstars, some polypores, jelly fungi, ergots, and smuts.
It is often the case that features of the stipe are required to make a positive identification of a mushroom.
Such distinguishing characters include: the texture of the stipe (fibrous, brittle, chalky, leathery, firm, etc.) whether it has remains of a partial veil (such as an annulus or cortina) or universal veil (volva) whether
the stipes of many mushrooms fuse at their baseits general size and shapewhether the stipe extends underground in a root-like structure (a rhizome)
When collecting mushrooms for identification it is critical to maintain all these characters intact by digging the mushroom out of the soil, rather than cutting it off mid-stipe.
Division: Ascomycota
Class: Sordariomycetes
Order: Hypocreales
Family: Hypocreaceae
Genus: Podostroma
Species: P. cornu-damae
Binomial name Podostroma cornu-damae
(Patouillard) Hongo & Izawa



ID confirmation : Podostroma cornu-damae
(Japanese: カエンタケ, Hepburn: kaentake)
poison fire coral .
Credit : Serena Lee
(Japanese: カエンタケ, Hepburn: kaentake)
poison fire coral .
Credit : Serena Lee
www.straitstimes.com/asia/australianz/killer-japanese-fungus-poison-fire-coral-found-in-australia
www.jcu.edu.au/news/releases/2019/october/deadly-fungus-found-in-far-north-queensland
www.jcu.edu.au/news/releases/2019/october/deadly-fungus-found-in-far-north-queensland


One of world’s deadliest fungi, the Poison Fire Coral, has been found in Far North Queensland for the first time, a long way from its usual home in the mountains of Japan and Korea.
James Cook University’s (JCU) Dr Matt Barrett confirmed the identity of the Poison Fire Coral fungus after local photographer Ray Palmer provided an image of a mystery fungus snapped in Redlynch in suburban Cairns.
The bright red Poison Fire Coral fruit bodies were found on tree roots and soil. Dr Barrett warned people to resist the urge to pick up the eye-catching fungus.
“If found, the fungus should not be touched, and definitely not eaten. Of the hundred or so toxic mushrooms that are known to researchers, this is the only one in which the toxins can be absorbed through the skin,” said Dr Barrett, a mycologist from the JCU Australian Tropical Herbarium (ATH) who specialises in the study of fungi.
“This record extends the distribution of the fungus considerably, and it may be even more widespread in tropical Australia,” said Dr Barrett.
Encyclopedia Britannica lists the species as the world’s second deadliest fungus. Several fatalities have been documented in Japan and Korea where people have brewed and drunk a tea with Poison Fire Coral, having confused it for the edible Ganoderma (Lingzhi or Reishi) or Cordyceps (vegetable caterpillar), which are used in traditional medicines.
The Poison Fire Coral produces at least eight toxic compounds that can be absorbed through the skin.
“Just touching the Fire Coral fungus can cause dermatitis (reddening or swelling of the skin). If eaten, it causes a horrifying array of symptoms: initially stomach pain, vomiting, diarrhea, fever and numbness, followed over hours or days by delamination of skin on face, hands and feet, and shrinking of the brain, which, in turn, causes altered perception, motion difficulties and speech impediments,” explained Dr Barrett.
If left untreated, death can occur from multiple organ failure or brain nerve dysfunction.
Dr Barrett said it was most likely that the fungus had occurred naturally in Cairns. There have also been reported instances of it growing in Papua New Guinea and Indonesia.
Scientists from ATH are re-evaluating the scientific name of this fungus in light of genetic and microscopic characteristics.
Dr Barrett joined ATH this year and since moving to Cairns from Western Australia, he has assisted many citizen scientists to put a name to their fungi finds and photos.
The Poison fire Coral is just one of more than 20 fungi species Dr Barrett has identified for local people that were not previously known to live in northern Queensland.
“The fact that we can find such a distinctive and medically important fungus like Poison Coral Fire Coral right in our backyard shows we have much to learn about fungi in northern Australia,” said Dr Barrett.
Contacts
Dr Matt Barrett (Cairns)
[email protected]
James Cook University’s (JCU) Dr Matt Barrett confirmed the identity of the Poison Fire Coral fungus after local photographer Ray Palmer provided an image of a mystery fungus snapped in Redlynch in suburban Cairns.
The bright red Poison Fire Coral fruit bodies were found on tree roots and soil. Dr Barrett warned people to resist the urge to pick up the eye-catching fungus.
“If found, the fungus should not be touched, and definitely not eaten. Of the hundred or so toxic mushrooms that are known to researchers, this is the only one in which the toxins can be absorbed through the skin,” said Dr Barrett, a mycologist from the JCU Australian Tropical Herbarium (ATH) who specialises in the study of fungi.
“This record extends the distribution of the fungus considerably, and it may be even more widespread in tropical Australia,” said Dr Barrett.
Encyclopedia Britannica lists the species as the world’s second deadliest fungus. Several fatalities have been documented in Japan and Korea where people have brewed and drunk a tea with Poison Fire Coral, having confused it for the edible Ganoderma (Lingzhi or Reishi) or Cordyceps (vegetable caterpillar), which are used in traditional medicines.
The Poison Fire Coral produces at least eight toxic compounds that can be absorbed through the skin.
“Just touching the Fire Coral fungus can cause dermatitis (reddening or swelling of the skin). If eaten, it causes a horrifying array of symptoms: initially stomach pain, vomiting, diarrhea, fever and numbness, followed over hours or days by delamination of skin on face, hands and feet, and shrinking of the brain, which, in turn, causes altered perception, motion difficulties and speech impediments,” explained Dr Barrett.
If left untreated, death can occur from multiple organ failure or brain nerve dysfunction.
Dr Barrett said it was most likely that the fungus had occurred naturally in Cairns. There have also been reported instances of it growing in Papua New Guinea and Indonesia.
Scientists from ATH are re-evaluating the scientific name of this fungus in light of genetic and microscopic characteristics.
Dr Barrett joined ATH this year and since moving to Cairns from Western Australia, he has assisted many citizen scientists to put a name to their fungi finds and photos.
The Poison fire Coral is just one of more than 20 fungi species Dr Barrett has identified for local people that were not previously known to live in northern Queensland.
“The fact that we can find such a distinctive and medically important fungus like Poison Coral Fire Coral right in our backyard shows we have much to learn about fungi in northern Australia,” said Dr Barrett.
Contacts
Dr Matt Barrett (Cairns)
[email protected]


Shot with torch


Shot with flash
Clavaria zollingeri
coral fungi
coral fungi

Clavaria zollingeri, commonly known as the violet coral or the magenta coral, is a widely distributed species of fungus.
It produces striking tubular, purple to pinkish-violet fruit bodies that grow up to 10 cm (3.9 in) tall and 7 cm (2.8 in) wide. The extreme tips of the fragile, slender branches are usually rounded and brownish.
A typical member of the clavarioid or club fungi,
Clavaria zollingeri is saprobic, and so derives nutrients by breaking down organic matter.
The fruit bodies are typically found growing on the ground in woodland litter, or in grasslands.
Variations in branching and color can often be used to distinguish C. zollingeri from similarly colored coral fungi such as Alloclavaria purpurea and Clavulina amethystina, although microscopy is required to reliably identify the latter species.
The coloring of the fruit bodies is quite variable, ranging from violet to amethyst, or violet shaded with brown or red. The colors may be variable over the fruit body; in one instance the outside branches were brown while the inner branches in the center of the bundle were light violet. Dried specimens may lose their coloring almost entirely, as the pigments may be sensitive to light or dryness.
The fruit body is typically 5 to 10 cm (2.0 to 3.9 in) tall and 4 to 7 cm (1.6 to 2.8 in) wide. The stem, or base, is short, and the branching starts a short distance above the ground. The surfaces of the fragile branches are smooth and dry; the branches are 2–6 thick, typically with rounded tips.
The fruit bodies of Clavaria zollingeri grow either solitarily, in groups, or in clusters on the ground in grassy spots, usually near hardwood trees, or with mosses.
It is a saprobic species, deriving nutrients by breaking down organic matter. It has a widespread distribution, and has been found in Australia, New Zealand, North America, South America, and Asia (including Brunei, India, and Korea.
Clavaria zollingeri contains lectins, a class of proteins that bind specific carbohydrates on the surface of cells, causing them to clump together.
A Korean study demonstrated that extracts of the fungus caused lymphoagglutination, a specific form of agglutination that involves white blood cells. In general, lectins are used in blood typing and serology, and they are widely used in affinity chromatography for purifying proteins.
Division: Basidiomycota
Class: Agaricomycetes
Order: Agaricales
Family: Clavariaceae
Genus: Clavaria
Species: C. zollingeri
Binomial name Clavaria zollingeri
Lév. (1846)
It produces striking tubular, purple to pinkish-violet fruit bodies that grow up to 10 cm (3.9 in) tall and 7 cm (2.8 in) wide. The extreme tips of the fragile, slender branches are usually rounded and brownish.
A typical member of the clavarioid or club fungi,
Clavaria zollingeri is saprobic, and so derives nutrients by breaking down organic matter.
The fruit bodies are typically found growing on the ground in woodland litter, or in grasslands.
Variations in branching and color can often be used to distinguish C. zollingeri from similarly colored coral fungi such as Alloclavaria purpurea and Clavulina amethystina, although microscopy is required to reliably identify the latter species.
The coloring of the fruit bodies is quite variable, ranging from violet to amethyst, or violet shaded with brown or red. The colors may be variable over the fruit body; in one instance the outside branches were brown while the inner branches in the center of the bundle were light violet. Dried specimens may lose their coloring almost entirely, as the pigments may be sensitive to light or dryness.
The fruit body is typically 5 to 10 cm (2.0 to 3.9 in) tall and 4 to 7 cm (1.6 to 2.8 in) wide. The stem, or base, is short, and the branching starts a short distance above the ground. The surfaces of the fragile branches are smooth and dry; the branches are 2–6 thick, typically with rounded tips.
The fruit bodies of Clavaria zollingeri grow either solitarily, in groups, or in clusters on the ground in grassy spots, usually near hardwood trees, or with mosses.
It is a saprobic species, deriving nutrients by breaking down organic matter. It has a widespread distribution, and has been found in Australia, New Zealand, North America, South America, and Asia (including Brunei, India, and Korea.
Clavaria zollingeri contains lectins, a class of proteins that bind specific carbohydrates on the surface of cells, causing them to clump together.
A Korean study demonstrated that extracts of the fungus caused lymphoagglutination, a specific form of agglutination that involves white blood cells. In general, lectins are used in blood typing and serology, and they are widely used in affinity chromatography for purifying proteins.
Division: Basidiomycota
Class: Agaricomycetes
Order: Agaricales
Family: Clavariaceae
Genus: Clavaria
Species: C. zollingeri
Binomial name Clavaria zollingeri
Lév. (1846)


Flavoparmelia caperata
greenshield lichen
greenshield lichen

Flavoparmelia caperata, the common greenshield lichen, is a foliose lichen that grows on the bark of trees, and occasionally on rock.
Flavoparmelia caperata' is a medium to large foliose lichen that has a very distinctive pale yellow green upper cortex when dry.
The rounded lobes, measuring 3–8 mm (0.1–0.3 in) wide, usually have patches of granular soredia arising from pustules. The lobes of the thallus may be smooth, but quite often have a wrinkled appearance especially in older specimens. The lower surface is black except for a brown margin; rhizoids attached to the lower surface are black and unbranched.
The very similar Flavoparmelia baltimorensis grows mainly on rock and has globose, pustular outgrowths (somewhat similar to isidia) on the upper surface of the lobes, but does not produce granular soredia.
Kingdom: Fungi
Division: Ascomycota
Class: Lecanoromycetes
Order: Lecanorales
Family: Parmeliaceae
Genus: Flavoparmelia
Species: F. caperata
Binomial name Flavoparmelia caperata
(L.) Hale (1986)
Flavoparmelia caperata' is a medium to large foliose lichen that has a very distinctive pale yellow green upper cortex when dry.
The rounded lobes, measuring 3–8 mm (0.1–0.3 in) wide, usually have patches of granular soredia arising from pustules. The lobes of the thallus may be smooth, but quite often have a wrinkled appearance especially in older specimens. The lower surface is black except for a brown margin; rhizoids attached to the lower surface are black and unbranched.
The very similar Flavoparmelia baltimorensis grows mainly on rock and has globose, pustular outgrowths (somewhat similar to isidia) on the upper surface of the lobes, but does not produce granular soredia.
Kingdom: Fungi
Division: Ascomycota
Class: Lecanoromycetes
Order: Lecanorales
Family: Parmeliaceae
Genus: Flavoparmelia
Species: F. caperata
Binomial name Flavoparmelia caperata
(L.) Hale (1986)


Ceratiomyxa fruticulosa
Slime mould sp
Slime mould sp

Ceratiomyxa fruticulosa is a slime mold. While not fungi, slime molds often form spore-bearing
structures that resemble those of the true fungi.
Although many slime mold species fruit on wood they do not form a penetrating and absorptive mass of
hyphae in the wood substrate.
Rather, slime molds form structures called
plasmodia which are naked (i.e., without cell walls) masses of protoplasm which can move and engulf
particles of food in an amoeboid manner.
Slime mold plasmodia creep about over the surfaces
of materials, engulfing bacteria, spores of fungi and
plants, protozoa, and particles of nonliving organic
matter. At some point, plasmodia convert into spore-bearing structures.
In Ceratiomyxa fruticulosa, the plasmodium
converts into a clustered mass of stalks bearing spores on their surfaces. There is evidence that the spores are actually one-celled sporangia.
Ceratiomyxa is a genus of plasmodial slime mould within the Eumycetozoa, first described by Pier Antonio Micheli. They are widely distributed and commonly found on decaying wood.
The plasmodium often appears as white frost-like growth or thin watery layers on wood. Pillar or wall-like sporangia bud from the plasmodium and develop spores that undergo multiple divisions before they release flagellated zoospores. The zoospores will then pair off, undergo plasmogamy, and form zygotes that will later form new plasmodia.
The genus currently contains 4 species. The most notable member is Ceratiomyxa fruticulosa, a slime mould found in most parts of the world. Other known species of Ceratiomyxa are mostly found in the tropics.
Ceratiomyxa comes from the Latin word ceratus meaning "waxed" and the ancient Greek word myxa meaning "mucus".
Ceratiomyxa was first described under the name Puccinia ramose (later revised to Ceratiomyxa fruticulosa) in 1729 by Pier Antonio Micheli, a pioneer of mycology.
In 1805, Albertini and Schweinitz described 2 more species in the genus, C. hydnoides and C. porioides. However, because Ceratiomyxa species can take a variety of different forms, many forms first described as species are later considered synonyms. This includes C. hydnoideum and C. porioides which are now often considered synonyms of C.fruticulosa.
Ceratiomyxa was first placed in the subclass Ceratiomyxomycetidae of the class Myxomycetes by Martin and Alexopoulos in 1969, but was later moved to subclass Protostelia of the class Eumycetozoa by Olive in 1970.
Ceratiomyxa is commonly found on rotting wood. Large logs and stumps are cited as ideal substrates for growth, although smaller colonies can also be found a tree branches. The Ceratiomyxa collection of Henry C. Gilbert has specimen growing on various evergreen coniferous trees (Pseudotsuga), elm (Ulmus), maple (Acer), oak (Quercus), Tilia, and willows (Salix). A specimen collected by F. O. Grover was found growing on a burlap sack.
Species of Ceratiomyxa can be found world-wide. C. fruticulosa is the most common species and has a cosmopolitan distribution. C. morchella and C.sphaerasperma have only been recorded in the tropics.
Plasmodium: Net-like or thin layer enveloped in mucous. Often translucent or white in colour but can also be tinted slightly yellow, pink, or blue-green.
The protoplasm within the plasmodium can be seen flowing, resting, and resuming flow in the opposite direction. This occurs in intervals of approximately 40 seconds. Protoplasmic flow slows as the sporangia develop and halt completely during mitosis.
Sporangia: Can be found in many different forms.
Common forms include, Simple pillar, Pillar-like
projections. In some species, pillars expand at the base and fuse to form a layer of plasmodium.
Fruticulus: Clusters of finger-like projections that are usually undivided.
Arbuscula: Branched resembling small trees. Spores develop on branch surfaces.
Filiforme: Long and slender pillars.
Porioides: Net-like formations of walls. Spores develop on sides and edges of wall.
Spores: The spores are round or oval globules individually attached to stalks sprouting from sporangia.
Spores vary in size (8-13 µ in diameter). The spore walls are thin and transparent. Spores appear grainy and may have some vacuoles which are often near the perimeter or surrounding the nucleus. The nucleus within the spore is about 3 µ in diameter.
Swarmcells: Oblong with either one long flagellum, one long and one short flagellum, or 2 long flagella. The flagella may occur together or at opposite ends of the cell. Upon emergence, the swarmcells are 6-8 µ in length and can develop to around 12 µ in older cells.
Ceratiomyxa plasmodia is diploid and can often be found emerging from rotting wood and spreading into a thin layer. The plasmodium then fruits to form sporangia, erect pillar or wall-like structures. The sporangia mature in about six hours. Many small stalks emerge from the sporangia to form protospores. Meiosis then begins in the protospores. Synapsis of the chromosomes in prophase is achieved at this stage.
After the stalks have fully elongated, a thin transparent wall is secreted around the protospores to become spores. Spores take approximately 24 hours to mature. The first meiotic division is then completed in the spores followed by a second meiotic division which leaves the spore with 4 nuclei.
After dispersal, these four-nucleate spores germinate and naked protoplasts emerge. The protoplasts take in water upon emergence and enlarge to about 3 times the size of the spore from which they emerged. The protoplasts then form short thread-like filaments that later retract back into round globules.
In some cases, the protoplast skips the thread phase and remains isodiametric. The nuclei then migrate to four points of a tetrahedron and the protoplast cleaves into a tetrad of lobes. Soon after, the tetrad undergoes mitosis and splits again into an octette of haploid cells.
Each cell releases a swarmcell with either one long flagellum, 2 unequal flagella, or 2 long flagella. The flagella are apical and whiplash type. The swarmcells may then lose their flagella and become asexual myxamoebae or undergo syngamy in pairs to produce a diploid zygote.
Domain: Eukaryota
Phylum: Amoebozoa
Class: Protostelia
Order: Protosteliida
Family: Ceratiomyxidae
Genus: Ceratiomyxa
J. Schrot. (1889)
Type species Ceratiomyxa mucida
(Pers.) J. Schröt.
structures that resemble those of the true fungi.
Although many slime mold species fruit on wood they do not form a penetrating and absorptive mass of
hyphae in the wood substrate.
Rather, slime molds form structures called
plasmodia which are naked (i.e., without cell walls) masses of protoplasm which can move and engulf
particles of food in an amoeboid manner.
Slime mold plasmodia creep about over the surfaces
of materials, engulfing bacteria, spores of fungi and
plants, protozoa, and particles of nonliving organic
matter. At some point, plasmodia convert into spore-bearing structures.
In Ceratiomyxa fruticulosa, the plasmodium
converts into a clustered mass of stalks bearing spores on their surfaces. There is evidence that the spores are actually one-celled sporangia.
Ceratiomyxa is a genus of plasmodial slime mould within the Eumycetozoa, first described by Pier Antonio Micheli. They are widely distributed and commonly found on decaying wood.
The plasmodium often appears as white frost-like growth or thin watery layers on wood. Pillar or wall-like sporangia bud from the plasmodium and develop spores that undergo multiple divisions before they release flagellated zoospores. The zoospores will then pair off, undergo plasmogamy, and form zygotes that will later form new plasmodia.
The genus currently contains 4 species. The most notable member is Ceratiomyxa fruticulosa, a slime mould found in most parts of the world. Other known species of Ceratiomyxa are mostly found in the tropics.
Ceratiomyxa comes from the Latin word ceratus meaning "waxed" and the ancient Greek word myxa meaning "mucus".
Ceratiomyxa was first described under the name Puccinia ramose (later revised to Ceratiomyxa fruticulosa) in 1729 by Pier Antonio Micheli, a pioneer of mycology.
In 1805, Albertini and Schweinitz described 2 more species in the genus, C. hydnoides and C. porioides. However, because Ceratiomyxa species can take a variety of different forms, many forms first described as species are later considered synonyms. This includes C. hydnoideum and C. porioides which are now often considered synonyms of C.fruticulosa.
Ceratiomyxa was first placed in the subclass Ceratiomyxomycetidae of the class Myxomycetes by Martin and Alexopoulos in 1969, but was later moved to subclass Protostelia of the class Eumycetozoa by Olive in 1970.
Ceratiomyxa is commonly found on rotting wood. Large logs and stumps are cited as ideal substrates for growth, although smaller colonies can also be found a tree branches. The Ceratiomyxa collection of Henry C. Gilbert has specimen growing on various evergreen coniferous trees (Pseudotsuga), elm (Ulmus), maple (Acer), oak (Quercus), Tilia, and willows (Salix). A specimen collected by F. O. Grover was found growing on a burlap sack.
Species of Ceratiomyxa can be found world-wide. C. fruticulosa is the most common species and has a cosmopolitan distribution. C. morchella and C.sphaerasperma have only been recorded in the tropics.
Plasmodium: Net-like or thin layer enveloped in mucous. Often translucent or white in colour but can also be tinted slightly yellow, pink, or blue-green.
The protoplasm within the plasmodium can be seen flowing, resting, and resuming flow in the opposite direction. This occurs in intervals of approximately 40 seconds. Protoplasmic flow slows as the sporangia develop and halt completely during mitosis.
Sporangia: Can be found in many different forms.
Common forms include, Simple pillar, Pillar-like
projections. In some species, pillars expand at the base and fuse to form a layer of plasmodium.
Fruticulus: Clusters of finger-like projections that are usually undivided.
Arbuscula: Branched resembling small trees. Spores develop on branch surfaces.
Filiforme: Long and slender pillars.
Porioides: Net-like formations of walls. Spores develop on sides and edges of wall.
Spores: The spores are round or oval globules individually attached to stalks sprouting from sporangia.
Spores vary in size (8-13 µ in diameter). The spore walls are thin and transparent. Spores appear grainy and may have some vacuoles which are often near the perimeter or surrounding the nucleus. The nucleus within the spore is about 3 µ in diameter.
Swarmcells: Oblong with either one long flagellum, one long and one short flagellum, or 2 long flagella. The flagella may occur together or at opposite ends of the cell. Upon emergence, the swarmcells are 6-8 µ in length and can develop to around 12 µ in older cells.
Ceratiomyxa plasmodia is diploid and can often be found emerging from rotting wood and spreading into a thin layer. The plasmodium then fruits to form sporangia, erect pillar or wall-like structures. The sporangia mature in about six hours. Many small stalks emerge from the sporangia to form protospores. Meiosis then begins in the protospores. Synapsis of the chromosomes in prophase is achieved at this stage.
After the stalks have fully elongated, a thin transparent wall is secreted around the protospores to become spores. Spores take approximately 24 hours to mature. The first meiotic division is then completed in the spores followed by a second meiotic division which leaves the spore with 4 nuclei.
After dispersal, these four-nucleate spores germinate and naked protoplasts emerge. The protoplasts take in water upon emergence and enlarge to about 3 times the size of the spore from which they emerged. The protoplasts then form short thread-like filaments that later retract back into round globules.
In some cases, the protoplast skips the thread phase and remains isodiametric. The nuclei then migrate to four points of a tetrahedron and the protoplast cleaves into a tetrad of lobes. Soon after, the tetrad undergoes mitosis and splits again into an octette of haploid cells.
Each cell releases a swarmcell with either one long flagellum, 2 unequal flagella, or 2 long flagella. The flagella are apical and whiplash type. The swarmcells may then lose their flagella and become asexual myxamoebae or undergo syngamy in pairs to produce a diploid zygote.
Domain: Eukaryota
Phylum: Amoebozoa
Class: Protostelia
Order: Protosteliida
Family: Ceratiomyxidae
Genus: Ceratiomyxa
J. Schrot. (1889)
Type species Ceratiomyxa mucida
(Pers.) J. Schröt.


Geastrum fimbriatum
Earthstar fungus
Earthstar fungus

Geastrales is an order of gasterocarpic basidiomycetes (fungi) that are related to Cantharellales. The order contains the single family Geastraceae, commonly known as "earthstars", which older classifications had placed in Lycoperdales, or Phallales.
Approximately 64 species are classified in this family, divided among eight genera, including the Geastrum, Myriostoma and Sphaerobolus.
The Sphaerobolus are known as "shotgun fungus" or "cannonball fungus". They colonize wood-based mulches and may throw black, sticky, spore-containing globs onto nearby surfaces.
The fruiting bodies of several earthstars are hygroscopic: in dry weather the "petals" will dry and curl up around the soft spore sac, protecting it. In this state, often the whole fungus becomes detached from the ground and may roll around like a tumbleweed. Once mature, their exoperidium splits into a variable number of rays, which give Geastrum their visible star shape. The exoperidial rays are there to protect the endoperidial body and orchestrate spore dispersal.
In wetter weather, the "petals" moisten and uncurl; some even curl backward lifting the spore sac up. This allows rain or animals to hit the spore sac so, emitting spores when enough moisture is present for them to germinate and establish.
Geastrum fimbriatum, commonly known as the fringed earthstar or the sessile earthstar, is an inedible species of mushroom belonging to the genus Geastrum, or earthstar fungi. First described in 1829, the species has a widespread distribution, and is found in Asia, Europe, and the Americas.
It is distinguished from other earthstars by the delicate fibers that line the circular pore at the top of its spore sac.
The fruit bodies of Geastrum fimbriatum start out roughly spherical and hypogeous. As it matures, it pushed up through the soil and the other layer of the spore case (exoperidium) splits open to form between 5 and 8 rays that curve downward.
The fully expanded fruit body has a diameter of up to 3 cm (1.2 in). Before expansion, the outer surface has a cottony surface with adherent soil particles; this ultimately peels off to reveal a smooth, grayish-brown surface.
The inner spore sac is yellowish brown and features a small conical pore with fringed edges. Unlike other similar earthstar fungi, the edges of this pore are not sharply delimited from the rest of the spore sac, and do not have grooves. The fruit bodies have no distinctive taste or odor.
The spores are spherical, roughened by many small points or warts, and measure 2.4–4 μm. The capillitium is thick-walled, unbranched, and 4–7 μm thick.
Similar species include Geastrum saccatum, which is larger – up to 5 cm (2.0 in) across – and has a clearly delimited ring-like area around the pore opening. Geastrum rufescens has reddish tones that are absent from G. fimbriatum.
Although typically listed by field guides as an inedible species, it is eaten by the tribal peoples of Madhya Pradesh.
Geastrum fimbriatum is a saprobic species, and it fruit bodies grow on the ground in groups or clusters, usually near the stumps of hardwood trees. It is found in Asia, India, Mongolia, eastern North Americal (including Mexico), Central America (Costa Rica), South America (Brazil), and Europe.
Division: Basidiomycota
Class: Agaricomycetes
Subclass: Phallomycetidae
Order: Geastrales
K.Hosaka & Castellano (2007)
Family: Geastraceae
Corda (1842)
Type genus Geastrum
Pers. (1801)
Species: G. fimbriatum
Binomial name Geastrum fimbriatum
Fr. (1829)
Approximately 64 species are classified in this family, divided among eight genera, including the Geastrum, Myriostoma and Sphaerobolus.
The Sphaerobolus are known as "shotgun fungus" or "cannonball fungus". They colonize wood-based mulches and may throw black, sticky, spore-containing globs onto nearby surfaces.
The fruiting bodies of several earthstars are hygroscopic: in dry weather the "petals" will dry and curl up around the soft spore sac, protecting it. In this state, often the whole fungus becomes detached from the ground and may roll around like a tumbleweed. Once mature, their exoperidium splits into a variable number of rays, which give Geastrum their visible star shape. The exoperidial rays are there to protect the endoperidial body and orchestrate spore dispersal.
In wetter weather, the "petals" moisten and uncurl; some even curl backward lifting the spore sac up. This allows rain or animals to hit the spore sac so, emitting spores when enough moisture is present for them to germinate and establish.
Geastrum fimbriatum, commonly known as the fringed earthstar or the sessile earthstar, is an inedible species of mushroom belonging to the genus Geastrum, or earthstar fungi. First described in 1829, the species has a widespread distribution, and is found in Asia, Europe, and the Americas.
It is distinguished from other earthstars by the delicate fibers that line the circular pore at the top of its spore sac.
The fruit bodies of Geastrum fimbriatum start out roughly spherical and hypogeous. As it matures, it pushed up through the soil and the other layer of the spore case (exoperidium) splits open to form between 5 and 8 rays that curve downward.
The fully expanded fruit body has a diameter of up to 3 cm (1.2 in). Before expansion, the outer surface has a cottony surface with adherent soil particles; this ultimately peels off to reveal a smooth, grayish-brown surface.
The inner spore sac is yellowish brown and features a small conical pore with fringed edges. Unlike other similar earthstar fungi, the edges of this pore are not sharply delimited from the rest of the spore sac, and do not have grooves. The fruit bodies have no distinctive taste or odor.
The spores are spherical, roughened by many small points or warts, and measure 2.4–4 μm. The capillitium is thick-walled, unbranched, and 4–7 μm thick.
Similar species include Geastrum saccatum, which is larger – up to 5 cm (2.0 in) across – and has a clearly delimited ring-like area around the pore opening. Geastrum rufescens has reddish tones that are absent from G. fimbriatum.
Although typically listed by field guides as an inedible species, it is eaten by the tribal peoples of Madhya Pradesh.
Geastrum fimbriatum is a saprobic species, and it fruit bodies grow on the ground in groups or clusters, usually near the stumps of hardwood trees. It is found in Asia, India, Mongolia, eastern North Americal (including Mexico), Central America (Costa Rica), South America (Brazil), and Europe.
Division: Basidiomycota
Class: Agaricomycetes
Subclass: Phallomycetidae
Order: Geastrales
K.Hosaka & Castellano (2007)
Family: Geastraceae
Corda (1842)
Type genus Geastrum
Pers. (1801)
Species: G. fimbriatum
Binomial name Geastrum fimbriatum
Fr. (1829)




Tremella foliacea

Tremella foliacea is a species of fungus producing brownish, frondose, gelatinous basidiocarps (fruit bodies).
It is widespread, particularly in north temperate regions, and is parasitic on other species of fungi (Stereum spp.), that grow on dead attached and recently fallen branches of broadleaf trees and conifers.
Common names include leafy brain, jelly leaf, and brown witch's butter. The species is said to be edible, but is not much valued. Fruit bodies are gelatinous, pale to dark brown, sometimes purple-brown to almost black, up to 20 cm (8 in) across, and seaweed-like (with branched, undulating fronds).
Microscopically, the hyphae are clamped and occur in a dense gelatinous matrix. Haustorial cells arise on the hyphae, producing filaments that attach to and penetrate the hyphae of the host. The basidia are tremelloid (globose to ellipsoid, with oblique to vertical septa), 10 to 19 by 8 to 14 μm, usually unstalked. The basidiospores are mostly ellipsoid, smooth, 6.5 to 10 by 4.5 to 8 μm, and germinate by hyphal tube or by yeast cells.
When fully developed Tremella foliacea is even more contorted than its close relatives Tremella mesenterica and Tremella aurantiaca. In dry weather these fungi shrivel to hard blackish crusts, in which state they are much more difficult to spot. When it rains the fruitbodies rehydrate and turn translucent again.
Late summer and autumn are the best times to look for Tremella foliacea, which is capable of sexual and asexual reproduction: it propagates not only via basidiospores but also by producing conidiospores.
Initially disc-like, the fruitbody soon develops irregular convoluted folds with rounded margins. Individual fruitbodies grow to between 2 and 20cm across. The leaf-like clusters of folded lobes are usually reddish-brown or pale pinkish-brown but sometimes reddish orange; gelatinous when damp, turning dark and shriveling to an inconspicuous fraction of its former size during very dry weather.
Leafy Brain fungus is fairly common throughout Britain and Ireland and is found also in other European countries from Norway right down to Portugal. This species is also recorded in most other temperate regions including North Africa, Asia, Australia and both North America and South America. It is truly a worldwide fungus.
This jelly fungus was originally described in 1800 by Christiaan Hendrik Persoon, who called it Tremella foliacea, by which name mycologists still refer to it today.
Tremella foliacea has many synonyms including Tremella frondosa, Tremella fimbriata Pers., Gyraria foliacea (Pers.) Gray, Tremella succinea Pers., Tremella nigrescens Fr., Ulocolla foliacea (Pers.) Bref., Exidia foliacea (Pers.) P. Karst., Phaeotremella pseudofoliacea Rea, and Tremella foliacea var. succinea (Pers.) Neuhof.
This jelly fungus grows on dead timber of broadleaf trees and conifers. It is not the dead timber that Tremella foliacea consumes but rather a kind of bracket or crust fungus that itself has been feeding on the wood. Tremella foliacea must therefore be - Classed as a parasitic rather than saprobic species.
This jelly-like fungus can be found throughout the year, but it is most visible in autumn and winter, young fruitbodies of Tremella foliacea are disc-like but they will become contorted and develop multiple folds as they expand and approach maturity.
Division: Basidiomycota
Class: Tremellomycetes
Order: Tremellales
Family: Tremellaceae
Genus: Tremella
Species: T. foliacea
Binomial name Tremella foliacea
Pers. (1800)
It is widespread, particularly in north temperate regions, and is parasitic on other species of fungi (Stereum spp.), that grow on dead attached and recently fallen branches of broadleaf trees and conifers.
Common names include leafy brain, jelly leaf, and brown witch's butter. The species is said to be edible, but is not much valued. Fruit bodies are gelatinous, pale to dark brown, sometimes purple-brown to almost black, up to 20 cm (8 in) across, and seaweed-like (with branched, undulating fronds).
Microscopically, the hyphae are clamped and occur in a dense gelatinous matrix. Haustorial cells arise on the hyphae, producing filaments that attach to and penetrate the hyphae of the host. The basidia are tremelloid (globose to ellipsoid, with oblique to vertical septa), 10 to 19 by 8 to 14 μm, usually unstalked. The basidiospores are mostly ellipsoid, smooth, 6.5 to 10 by 4.5 to 8 μm, and germinate by hyphal tube or by yeast cells.
When fully developed Tremella foliacea is even more contorted than its close relatives Tremella mesenterica and Tremella aurantiaca. In dry weather these fungi shrivel to hard blackish crusts, in which state they are much more difficult to spot. When it rains the fruitbodies rehydrate and turn translucent again.
Late summer and autumn are the best times to look for Tremella foliacea, which is capable of sexual and asexual reproduction: it propagates not only via basidiospores but also by producing conidiospores.
Initially disc-like, the fruitbody soon develops irregular convoluted folds with rounded margins. Individual fruitbodies grow to between 2 and 20cm across. The leaf-like clusters of folded lobes are usually reddish-brown or pale pinkish-brown but sometimes reddish orange; gelatinous when damp, turning dark and shriveling to an inconspicuous fraction of its former size during very dry weather.
Leafy Brain fungus is fairly common throughout Britain and Ireland and is found also in other European countries from Norway right down to Portugal. This species is also recorded in most other temperate regions including North Africa, Asia, Australia and both North America and South America. It is truly a worldwide fungus.
This jelly fungus was originally described in 1800 by Christiaan Hendrik Persoon, who called it Tremella foliacea, by which name mycologists still refer to it today.
Tremella foliacea has many synonyms including Tremella frondosa, Tremella fimbriata Pers., Gyraria foliacea (Pers.) Gray, Tremella succinea Pers., Tremella nigrescens Fr., Ulocolla foliacea (Pers.) Bref., Exidia foliacea (Pers.) P. Karst., Phaeotremella pseudofoliacea Rea, and Tremella foliacea var. succinea (Pers.) Neuhof.
This jelly fungus grows on dead timber of broadleaf trees and conifers. It is not the dead timber that Tremella foliacea consumes but rather a kind of bracket or crust fungus that itself has been feeding on the wood. Tremella foliacea must therefore be - Classed as a parasitic rather than saprobic species.
This jelly-like fungus can be found throughout the year, but it is most visible in autumn and winter, young fruitbodies of Tremella foliacea are disc-like but they will become contorted and develop multiple folds as they expand and approach maturity.
Division: Basidiomycota
Class: Tremellomycetes
Order: Tremellales
Family: Tremellaceae
Genus: Tremella
Species: T. foliacea
Binomial name Tremella foliacea
Pers. (1800)

Neonothopanus nambi
bioluminescence fungi
bioluminescence fungi

Neonothopanus is a genus of three species of fungi in the agaric family Marasmiaceae. The genus was circumscribed in 1999.
The type species N. nambi is found in Australia, South America, Central America, Singapore and Malaysia, while N. gardneri is found in South America.
Both of these species are bioluminescent.
N. hygrophanus, found in central Africa, was added to the genus in 2011.
N.nambi emits a faint white luminance when see in total darkness and one will by pass it unknowingly that this species are bioluminescent, compare to M.chlorophos it emits a strong green luminance in darkness and one will definately see it even from a distance.
N. nambi is a poisonous bioluminescent mushroom in the family Marasmiaceae. The genetic and molecular mechanisms underlying this species' bioluminescence were published in 2019, the first to be elucidated for a fungus. In 2020, genes from this fungus were used to create bioluminescent tobacco plants.
Kingdom: Fungi
Phylum: Basidiomycota
Class: Agaricomycetes
Subclass: Agaricomycetidae
Order: Agaricales
Family: Marasmiaceae
Genus: Neonothopanus
Species: N. nambi
Binomial name Neonothopanus nambi
(Speg.) R.H. Petersen & Krisai, 1999
The type species N. nambi is found in Australia, South America, Central America, Singapore and Malaysia, while N. gardneri is found in South America.
Both of these species are bioluminescent.
N. hygrophanus, found in central Africa, was added to the genus in 2011.
N.nambi emits a faint white luminance when see in total darkness and one will by pass it unknowingly that this species are bioluminescent, compare to M.chlorophos it emits a strong green luminance in darkness and one will definately see it even from a distance.
N. nambi is a poisonous bioluminescent mushroom in the family Marasmiaceae. The genetic and molecular mechanisms underlying this species' bioluminescence were published in 2019, the first to be elucidated for a fungus. In 2020, genes from this fungus were used to create bioluminescent tobacco plants.
Kingdom: Fungi
Phylum: Basidiomycota
Class: Agaricomycetes
Subclass: Agaricomycetidae
Order: Agaricales
Family: Marasmiaceae
Genus: Neonothopanus
Species: N. nambi
Binomial name Neonothopanus nambi
(Speg.) R.H. Petersen & Krisai, 1999
How is bioluminescence form
Bioluminescence is the production and emission of light by a living organism. It is a form of chemiluminescence.
Bioluminescence occurs widely in marine vertebrates and invertebrates, as well as in some fungi, microorganisms including some bioluminescent bacteria, and terrestrial arthropods such as fireflies.
Bioluminescence is used by living things to hunt prey, defend against predators, find mates, and execute other vital activities and some species luminesce to confuse attackers.
Most bioluminescent organisms are found in the ocean. These bioluminescent marine species include fish, bacteria, and jellies and Some bioluminescent organisms, including fireflies and fungi, are found on land. There are almost no bioluminescent organisms native to freshwater habitats.
In some animals, the light is bacteriogenic, produced by symbiotic bacteria such as those from the genus Vibrio; in others, it is autogenic, produced by the animals themselves.
Bioluminescence is a "cold light." Cold light means less than 20% of the light generates thermal radiation, or heat.
Luciferin is the compound that actually produces light. Luciferase is an enzyme. An enzyme is a chemical (called a catalyst) that interacts with a substrate to affect the rate of a chemical reaction.
In a general sense, the principal chemical reaction in bioluminescence involves a light-emitting molecule and an enzyme, generally called luciferin and luciferase, respectively.
In a chemical reaction, luciferin is called the substrate. The bioluminescent color (yellow in fireflies, greenish in lanternfish) is a result of the arrangement of luciferin molecules.
Some bioluminescent organisms produce (synthesize) luciferin on their own and some bioluminescent organisms do not synthesize luciferin. Instead, they absorb it through other organisms, either as food or in a symbiotic relationship.
The interaction of the luciferase with oxidized (oxygen-added) luciferin creates a byproduct, called oxyluciferin which is a type of chemical reaction creates light.
Most bioluminescent reactions involve luciferin and luciferase. Some reactions, however, do not involve an enzyme (luciferase). These reactions involve a chemical called a photoprotein. Photoproteins combine with luciferins and oxygen, but need another agent, often an ion of the element calcium, to produce light.
The appearance of bioluminescent light varies greatly, depending on the habitat and organism in which it is found.
Some species of fungi present in decaying wood, for instance, emit a fairly consistent glow, called foxfire and some organisms emit very bright light continuously.
Bioluminescence occurs widely in marine vertebrates and invertebrates, as well as in some fungi, microorganisms including some bioluminescent bacteria, and terrestrial arthropods such as fireflies.
Bioluminescence is used by living things to hunt prey, defend against predators, find mates, and execute other vital activities and some species luminesce to confuse attackers.
Most bioluminescent organisms are found in the ocean. These bioluminescent marine species include fish, bacteria, and jellies and Some bioluminescent organisms, including fireflies and fungi, are found on land. There are almost no bioluminescent organisms native to freshwater habitats.
In some animals, the light is bacteriogenic, produced by symbiotic bacteria such as those from the genus Vibrio; in others, it is autogenic, produced by the animals themselves.
Bioluminescence is a "cold light." Cold light means less than 20% of the light generates thermal radiation, or heat.
Luciferin is the compound that actually produces light. Luciferase is an enzyme. An enzyme is a chemical (called a catalyst) that interacts with a substrate to affect the rate of a chemical reaction.
In a general sense, the principal chemical reaction in bioluminescence involves a light-emitting molecule and an enzyme, generally called luciferin and luciferase, respectively.
In a chemical reaction, luciferin is called the substrate. The bioluminescent color (yellow in fireflies, greenish in lanternfish) is a result of the arrangement of luciferin molecules.
Some bioluminescent organisms produce (synthesize) luciferin on their own and some bioluminescent organisms do not synthesize luciferin. Instead, they absorb it through other organisms, either as food or in a symbiotic relationship.
The interaction of the luciferase with oxidized (oxygen-added) luciferin creates a byproduct, called oxyluciferin which is a type of chemical reaction creates light.
Most bioluminescent reactions involve luciferin and luciferase. Some reactions, however, do not involve an enzyme (luciferase). These reactions involve a chemical called a photoprotein. Photoproteins combine with luciferins and oxygen, but need another agent, often an ion of the element calcium, to produce light.
The appearance of bioluminescent light varies greatly, depending on the habitat and organism in which it is found.
Some species of fungi present in decaying wood, for instance, emit a fairly consistent glow, called foxfire and some organisms emit very bright light continuously.
Why Do Some Mushrooms Glow In The Dark?
Why Do Some Mushrooms Glow In The Dark?
Glowing in the dark costs energy.
The Secret Behind Bioluminescent Mushrooms’ Magic Glow.
Scientists use chemistry to account for an astonishing phenomenon. Researchers described the compound that gives the mushrooms their glow — It’s called oxyluciferin.
Mushrooms use luciferins—light-emitting compounds found in other glowing animals and plants—to attract insects. The bugs then help spread their spores to sheltered places in the forest, which helps the mushroom species survive.
Luciferins give fireflies and even bioluminescent underwater creatures their glow. Paired with an enzyme and oxygen, it releases light that illuminates the fungi.
Scientists went foraging for the glow-in-the-dark mushrooms and they crushed the mushrooms to make a slurry filled with luciferins. Then they isolated the luciferin and studied it, capturing its chemical structure and experimenting with its ability to fuel those flourescent colors.
Not only does the team now know that the mushrooms are fueled by their own kind of luciferin, but they also figured out that the enzyme that combines with the chemical to trigger light could be what they call “promiscuous.”
That means that the enzyme might be able to interact with different luciferins—and produce even more shades of that pretty glow. And that suggests that when it comes to these magical mushrooms, there’s even more to discover.
Fungi produce tiny spores to spread themselves, much like seeds from trees. Insects can help transport the fungal spores. Fruit bodies are produced to disperse their sexual progeny as spores. Many fungi shoot spores into the air from the underside of the mushrooms, relying on moving air currents to passively distribute the spores over a wide area.
Through aroma. Truffles, the fruiting body of the Ascomycete fungi, use their smell to attract fungivores such as pigs or squirrels who eat them and leave spores behind in their waste. Stinkhorn mushrooms have a foul-smelling slime which attracts flies and other insects. The flies eat the slime and unwittingly spread the spores elsewhere.
Since bioluminescence is so energetically costly, the mushrooms only glow at night. They don’t glow bright enough during the day to be extra visible. An added bonus is that spores prefer to become active and grow at night when it is more humid.
Bioluminescence is one of the factors attracting insects to the mushrooms to help them spread their spores.
"On a totally dark night, without any moon, if you have your light off, these green mushrooms are basically the only light source you see in the forest besides the fireflies."
You just have to turn off your flashlight and the mushrooms stand out if they're there.
Glowing in the dark costs energy.
The Secret Behind Bioluminescent Mushrooms’ Magic Glow.
Scientists use chemistry to account for an astonishing phenomenon. Researchers described the compound that gives the mushrooms their glow — It’s called oxyluciferin.
Mushrooms use luciferins—light-emitting compounds found in other glowing animals and plants—to attract insects. The bugs then help spread their spores to sheltered places in the forest, which helps the mushroom species survive.
Luciferins give fireflies and even bioluminescent underwater creatures their glow. Paired with an enzyme and oxygen, it releases light that illuminates the fungi.
Scientists went foraging for the glow-in-the-dark mushrooms and they crushed the mushrooms to make a slurry filled with luciferins. Then they isolated the luciferin and studied it, capturing its chemical structure and experimenting with its ability to fuel those flourescent colors.
Not only does the team now know that the mushrooms are fueled by their own kind of luciferin, but they also figured out that the enzyme that combines with the chemical to trigger light could be what they call “promiscuous.”
That means that the enzyme might be able to interact with different luciferins—and produce even more shades of that pretty glow. And that suggests that when it comes to these magical mushrooms, there’s even more to discover.
Fungi produce tiny spores to spread themselves, much like seeds from trees. Insects can help transport the fungal spores. Fruit bodies are produced to disperse their sexual progeny as spores. Many fungi shoot spores into the air from the underside of the mushrooms, relying on moving air currents to passively distribute the spores over a wide area.
Through aroma. Truffles, the fruiting body of the Ascomycete fungi, use their smell to attract fungivores such as pigs or squirrels who eat them and leave spores behind in their waste. Stinkhorn mushrooms have a foul-smelling slime which attracts flies and other insects. The flies eat the slime and unwittingly spread the spores elsewhere.
Since bioluminescence is so energetically costly, the mushrooms only glow at night. They don’t glow bright enough during the day to be extra visible. An added bonus is that spores prefer to become active and grow at night when it is more humid.
Bioluminescence is one of the factors attracting insects to the mushrooms to help them spread their spores.
"On a totally dark night, without any moon, if you have your light off, these green mushrooms are basically the only light source you see in the forest besides the fireflies."
You just have to turn off your flashlight and the mushrooms stand out if they're there.


Arcyria obvelata
Slime mold
Slime mold

Arcyria is a genus of Amoebozoa in the family Arcyriaceae. It includes the species Arcyria ferruginea.
Slime mould (Arcyria obvelata ), a tight group with sporangia fully expanded. the sporocarps of this slime mould are Sporangia, stalked, clustered and cylindrical in shape appearing rather finger like. In this case the spores have already been dispersed acquiring a rather spongy appearance. Also known as A. nutans.
Growth habit: gregarious to crowded, on rotting logs and stem of trees and tree rootings.
Amoebozoa is a major taxonomic group containing about 2,400 described species of amoeboid protists, often possessing blunt, fingerlike, lobose pseudopods and tubular mitochondrial cristae.
In most classification schemes, Amoebozoa is ranked as a phylum within either the kingdom Protista or the kingdom Protozoa. In the classification favored by the International Society of Protistologists, it is retained as an unranked "supergroup" within Eukaryota.
Molecular genetic analysis supports Amoebozoa as a monophyletic clade. Most phylogenetic trees identify it as the sister group to Opisthokonta, another major clade which contains both fungi and animals as well as some 300 species of unicellular protists.
Amoebozoa and Opisthokonta are sometimes grouped together in a high-level taxon, variously named Unikonta Amorphea or Opimoda.
Amoebozoa includes many of the best-known amoeboid organisms, such as Chaos, Entamoeba, Pelomyxa and the genus Amoeba itself. Species of Amoebozoa may be either shelled (testate) or naked, and cells may possess flagella.
Free-living species are common in both salt and freshwater as well as soil, moss and leaf litter. Some live as parasites or symbionts of other organisms, and some are known to cause disease in humans and other organisms.
While the majority of amoebozoan species are unicellular, the group also includes several clades of slime molds, which have a macroscopic, multicellular stage of life during which individual amoeboid cells remain together after multiple cell division to form a macroscopic plasmodium or, in cellular slime molds, aggregate to form one.
Amoebozoa vary greatly in size. Some are only 10–20 μm in diameter, while others are among the largest protozoa. The well-known species Amoeba proteus, which may reach 800 μm in length, is often studied in schools and laboratories as a representative cell or model organism, partly because of its convenient size.
Multinucleate amoebae like Chaos and Pelomyxa
may be several millimetres in length, and some multicellular amoebozoa, such as the "dog vomit" slime mold Fuligo septica, can cover an area of several square meters.
Amoebozoa is a large and diverse group, but certain features are common to many of its members. The amoebozoan cell is typically divided into a granular central mass, called endoplasm, and a clear outer layer, called ectoplasm. During locomotion, the endoplasm flows forwards and the ectoplasm runs backwards along the outside of the cell.
In motion, many amoebozoans have a clearly defined anterior and posterior and may assume a "monopodial" form, with the entire cell functioning as a single pseudopod. Large pseudopods may produce numerous clear projections called subpseudopodia (or determinate pseudopodia), which are extended to a certain length and then retracted, either for the purpose of locomotion or food intake.
A cell may also form multiple indeterminate pseudopodia, through which the entire contents of the cell flow in the direction of locomotion. These are more or less tubular and are mostly filled with granular endoplasm. The cell mass flows into a leading pseudopod, and the others ultimately retract, unless the organism changes direction.
While most amoebozoans are "naked," like the familiar Amoeba and Chaos, or covered with a loose coat of minute scales, like Cochliopodium and Korotnevella,
members of the order Arcellinida form rigid shells, or tests, equipped with a single aperture through which the pseudopods emerge. Arcellinid tests may be secreted from organic materials, as in Arcella, or built up from collected particles cemented together, as in Difflugia.
In all amoebozoa, the primary mode of nutrition is phagocytosis, in which the cell surrounds potential food particles with its pseudopods, sealing them into vacuoles within which they may be digested and absorbed.
Some amoebozoans have a posterior bulb called a uroid, which may serve to accumulate waste, periodically detaching from the rest of the cell. When food is scarce, most species can form cysts, which may be carried aerially and introduce them to new environments. In slime moulds, these structures are called spores, and form on stalked structures called fruiting bodies or sporangia.
The majority of Amoebozoa lack flagella and more generally do not form microtubule-supported structures except during mitosis. However, flagella do occur among the Archamoebae, and many slime moulds produce biflagellate gametes.
The flagellum is generally anchored by a cone of microtubules, suggesting a close relationship to the opisthokonts. The mitochondria in amoebozoan cells characteristically have branching tubular cristae. However, among the Archamoebae, which are adapted to anoxic or microaerophilic habitats, mitochondria have been lost.
Scientific classification
(unranked): Amoebozoa
Infraphylum: Mycetozoa
Class: Myxogastria
Order: Trichiida
Family: Arcyriaceae
Species: A. obvelata
Genus: Arcyria
Wiggers
Slime mould (Arcyria obvelata ), a tight group with sporangia fully expanded. the sporocarps of this slime mould are Sporangia, stalked, clustered and cylindrical in shape appearing rather finger like. In this case the spores have already been dispersed acquiring a rather spongy appearance. Also known as A. nutans.
Growth habit: gregarious to crowded, on rotting logs and stem of trees and tree rootings.
Amoebozoa is a major taxonomic group containing about 2,400 described species of amoeboid protists, often possessing blunt, fingerlike, lobose pseudopods and tubular mitochondrial cristae.
In most classification schemes, Amoebozoa is ranked as a phylum within either the kingdom Protista or the kingdom Protozoa. In the classification favored by the International Society of Protistologists, it is retained as an unranked "supergroup" within Eukaryota.
Molecular genetic analysis supports Amoebozoa as a monophyletic clade. Most phylogenetic trees identify it as the sister group to Opisthokonta, another major clade which contains both fungi and animals as well as some 300 species of unicellular protists.
Amoebozoa and Opisthokonta are sometimes grouped together in a high-level taxon, variously named Unikonta Amorphea or Opimoda.
Amoebozoa includes many of the best-known amoeboid organisms, such as Chaos, Entamoeba, Pelomyxa and the genus Amoeba itself. Species of Amoebozoa may be either shelled (testate) or naked, and cells may possess flagella.
Free-living species are common in both salt and freshwater as well as soil, moss and leaf litter. Some live as parasites or symbionts of other organisms, and some are known to cause disease in humans and other organisms.
While the majority of amoebozoan species are unicellular, the group also includes several clades of slime molds, which have a macroscopic, multicellular stage of life during which individual amoeboid cells remain together after multiple cell division to form a macroscopic plasmodium or, in cellular slime molds, aggregate to form one.
Amoebozoa vary greatly in size. Some are only 10–20 μm in diameter, while others are among the largest protozoa. The well-known species Amoeba proteus, which may reach 800 μm in length, is often studied in schools and laboratories as a representative cell or model organism, partly because of its convenient size.
Multinucleate amoebae like Chaos and Pelomyxa
may be several millimetres in length, and some multicellular amoebozoa, such as the "dog vomit" slime mold Fuligo septica, can cover an area of several square meters.
Amoebozoa is a large and diverse group, but certain features are common to many of its members. The amoebozoan cell is typically divided into a granular central mass, called endoplasm, and a clear outer layer, called ectoplasm. During locomotion, the endoplasm flows forwards and the ectoplasm runs backwards along the outside of the cell.
In motion, many amoebozoans have a clearly defined anterior and posterior and may assume a "monopodial" form, with the entire cell functioning as a single pseudopod. Large pseudopods may produce numerous clear projections called subpseudopodia (or determinate pseudopodia), which are extended to a certain length and then retracted, either for the purpose of locomotion or food intake.
A cell may also form multiple indeterminate pseudopodia, through which the entire contents of the cell flow in the direction of locomotion. These are more or less tubular and are mostly filled with granular endoplasm. The cell mass flows into a leading pseudopod, and the others ultimately retract, unless the organism changes direction.
While most amoebozoans are "naked," like the familiar Amoeba and Chaos, or covered with a loose coat of minute scales, like Cochliopodium and Korotnevella,
members of the order Arcellinida form rigid shells, or tests, equipped with a single aperture through which the pseudopods emerge. Arcellinid tests may be secreted from organic materials, as in Arcella, or built up from collected particles cemented together, as in Difflugia.
In all amoebozoa, the primary mode of nutrition is phagocytosis, in which the cell surrounds potential food particles with its pseudopods, sealing them into vacuoles within which they may be digested and absorbed.
Some amoebozoans have a posterior bulb called a uroid, which may serve to accumulate waste, periodically detaching from the rest of the cell. When food is scarce, most species can form cysts, which may be carried aerially and introduce them to new environments. In slime moulds, these structures are called spores, and form on stalked structures called fruiting bodies or sporangia.
The majority of Amoebozoa lack flagella and more generally do not form microtubule-supported structures except during mitosis. However, flagella do occur among the Archamoebae, and many slime moulds produce biflagellate gametes.
The flagellum is generally anchored by a cone of microtubules, suggesting a close relationship to the opisthokonts. The mitochondria in amoebozoan cells characteristically have branching tubular cristae. However, among the Archamoebae, which are adapted to anoxic or microaerophilic habitats, mitochondria have been lost.
Scientific classification
(unranked): Amoebozoa
Infraphylum: Mycetozoa
Class: Myxogastria
Order: Trichiida
Family: Arcyriaceae
Species: A. obvelata
Genus: Arcyria
Wiggers


The two photos above shows the spores releasing in Arcyria obvelata.

Amauroderma rugosum

Amauroderma is a genus of polypore fungi in the family Ganodermataceae. The genus, widespread in tropical areas, contains about 70 species.
Amauroderma fungi are wood-decay fungi that feed and fruit on decayed branches and trunks.
The fruit bodies of Amauroderma fungi comprise a cap and a stipe, and are typically woody, leathery, or corky in texture. The spores produced are usually spherical or nearly so, with a characteristic double wall structure that features U-shaped thickenings.
The fruit bodies of Amauroderma species are stipitate except in A. andina and may attain various shapes although centrally stipitate basidiocarps are most common. Several stipes may arise from the same base, frequently resulting in fused caps and compound fruit bodies. In section some fruit bodies are distinct with one or two distinct inner black bands or zones.
The stipe is often duplex with an outer dense layer surrounding an inner softer or hollow core sometimes separated by a black band. In species with a distinct tomentum on the stipe, there is often a dark zone just below the tomentum of the cap. These zones are absent from some species with a pale stipe without a tomentum. However, when present they continue into the context and frequently there is also another zone stretching more or less horizontally across the context.
Most basidiospores of Amauroderma mushrooms have an inner ornamented wall on which there is a hyaline (translucent) epicutis, which is very thin and difficult to see in ordinary microscopic preparations. Mature basidiospores are pale-yellowish. An apiculus
(a depressed area where the spore was once attached to the basidium via the sterigma) is often difficult to observe. Amauroderma is widespread in tropical areas.
Habitat loss from deforestation for oil palm plantation in tropical forests and partly collecting by local communities especially in Malaysia for medical properties. In Southeast Asia, more than 50% of Malaysia and Indonesia oil palm plantations in 2005 were on land that was forest in 1990 (Koh and Wilcove, 2008). Modification of substrates due to habitat loss will reduce population of species.
Amauroderma rugosum is a normally widespread in other regions but has been evaluated as a least concern (LC). This wild medicinal fungus always be consumed by indigenous communities in some countries.
Species can be found in tropical and subtropical areas. Collecting of this wild medicine fungus by human (Indigenous communities) and deforestation of diterocarp trees widely will reduce the population of fungus and degrade their natural habitat particulary in tropical rainforest regions in Thailand, Malaysia, Indonesia and Papua New Guinea
Population of species worldwide are facing habitat homogenization due to human activities especially in Asia. A low population density for this species in most regions due to extremely unfavorable habitats (substrate unfavourable for this saprophyte fungus) and habitat quality reduction then the species shows declines in some countries. Unclear of current trends of this species can be observed in most regions.
The species is growing as saprophytic (wood-decay fungi) on the ground with other plants and common in lowland dipterocarp of tropic forest and sub-tropic forest. The most suitable elevation for the growth and development of Amauroderma rugosum within on elevation of 200-1400m a.s.l. The higher humidity than 90% is the best for the growth and development of this species (Nguyen & Khanh, 2017).
In 2011 Conservation International listed the top 10 most endangered forests, characterized by having all lost 90% or more of their original habitat. Remaining habitat of endangered forests in Indo-Burma, New Caledonia, Sundaland, Philippines, Atlantic Forest, Mountains of Southwest China, California Floristic Province, Coastal Forests of Eastern Africa, Madagascar & Indian Ocean Islands and Eastern Afromontane only left between 11% and 5%.
Amauroderma rugosum is a wild mushroom that is worn as a necklace by the indigenous communities in Malaysia to prevent fits and incessant crying by babies.
Amauroderma rugosum, commonly known as “Jiǎzī” in China, is a wild mushroom traditionally used by the Chinese to reduce inflammation, to treat diuretic and upset stomach, and to prevent cancer.
It is also used by the indigenous communities in Malaysia to prevent epileptic episodes and incessant crying by babies. The aim of this study was to compare the wild and domesticated basidiocarps of A. rugosum for antioxidant and in vitro anti-inflammatory effects in LPS-stimulated RAW264.7 cells.
The wild basidiocarps of A. rugosum were collected from the Belum Forest, Perak, Malaysia and the domesticated basidiocarps of A. rugosum were cultivated in the mushroom house located in the University of Malaya, Kuala Lumpur, Malaysia. Both the wild and domesticated basidiocarps were subjected to ethanolic extraction and the extracts were tested for antioxidant and anti-inflammatory activities.
Amauroderma rugosum (AR) is a dietary mushroom in the Ganodermataceae family whose pharmacological activity and medicinal value have rarely been reported.
The antioxidant capacity and neuroprotective effects of AR were investigated. The aqueous extract of AR was confirmed to contain phenolic compounds, polysaccharides, and triterpenes. The results of 2,2-diphenyl-1-picryl-hydrazyl-hydrate (DPPH) and total antioxidant capacity assays revealed that AR extract scavenged reactive oxygen species. Moreover, AR extract decreased the cytotoxicity, oxidative stress, mitochondrial dysfunction, and apoptosis of PC12 cells induced by 6-hydroxydopamine (6-OHDA).
In addition, 6-OHDA upregulated the expressions of proapoptotic proteins and downregulated the Akt (protein kinase B)/mTOR- (mammalian target of rapamycin-) and MEK (mitogen-activated protein kinase kinase)/ERK- (extracellular signal-regulated kinases-) dependent signaling pathways. These effects of 6-OHDA were abolished or partially reversed by AR extract. Furthermore, the neuroprotective effects of AR in 6-OHDA-treated PC12 cells were significantly abolished by Akt and MEK inhibitor. Thus, AR extract possesses neuroprotective effects, probably through its antioxidant and antiapoptotic effects. These findings suggest the potential application of AR in the prevention or treatment of oxidative stress-related neurodegenerative diseases such as Parkinson’s disease.
To date, Parkinson’s disease remains the second most common neurodegenerative disease worldwide, and its incidence is increasing in people over 60 years of age. It is characterized by a selective and progressive loss of dopaminergic neurons in the substantia nigra pars compacta, leading to serious movement disturbances including postural instability, uncontrollable tremors, rigidity, and bradykinesia.
Currently, available treatments for Parkinson’s disease include dopaminergic replacement therapy and deep brain stimulation therapy. However, neither of these treatments can halt nor slow the progression of Parkinson’s disease. Therefore, it is crucial to develop novel drugs that can slow the neurodegenerative process.
Although the pathological mechanisms of Parkinson’s disease remain elusive, accumulating scientific evidence suggests that oxidative stress-induced cell injury plays an indispensable role in the degeneration of dopaminergic neurons. Interrupting the physiological maintenance of redox potential severely interferes with many biological processes in neurons, eventually leading to cell apoptosis.
Oxidative stress occurs when the rate of reactive oxygen species (ROS) scavenging is overwhelmed by the rate of ROS production. Excessive accumulation of ROS in dopaminergic neurons can damage most biological molecules, including lipids, proteins, and nucleic acids, thereby activating the intracellular inflammatory response, which induces cellular damage, mitochondrial dysfunction, oxidative DNA injury, and neuroinflammation. Therefore, decreasing oxidative injury in dopaminergic neurons has been widely proposed as an effective approach for the treatment of Parkinson’s disease.
Various experimental models have been established for investigating the role of oxidative stress in dopaminergic neuronal degeneration. These models involve the use of neurotoxins such as 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP), rotenone, 1,1-dimethyl-4,4-bipyridinium dichloride (paraquat) and 6-hydroxydopamine (6-OHDA).
The compound 6-OHDA, the hydroxylated analog of natural neurotransmitter dopamine, is a widely used neurotoxin that can be applied to establish different in vitro and in vivo Parkinson disease models. It is taken up by dopaminergic neurons via dopamine and norepinephrine transporters and is subsequently oxidized intracellularly, thereby releasing ROS including hydrogen peroxide, superoxide, and hydroxyl radicals.
Amauroderma rugosum (AR) is a basidiomycete in the Ganodermataceae family. This mushroom has a black stipe and a white surface covered with numerous pores. A notable characteristic of the mushroom is that its surface becomes red when it is scratched. Hence, it is also known as “blood Linzhi” in Chinese. Although AR is commonly consumed by people in China and South Asia, very few scientific studies have explored its beneficial effects on health or its medicinal value.
Nevertheless, a previous study reported that the extract of AR mycelia has antioxidant and anti-inflammatory effects in lipopolysaccharide-stimulated RAW 264.7 cells. Thus, AR might be a potential therapeutic agent or health supplement useful in the management of oxidative stress-related diseases. The aims of this study were to investigate the antioxidant capacity and neuroprotective activity of AR in a 6-OHDA-induced neurodegenerative cell model and to elucidate its underlying mechanisms of action.
Division: Basidiomycota
Class: Agaricomycetes
Order: Polyporales
Family: Ganodermataceae
Genus: Amauroderma
Murrill (1905)
Type species Amauroderma regulicolor
(Berk. ex Cooke) Murrill (1905)
Amauroderma fungi are wood-decay fungi that feed and fruit on decayed branches and trunks.
The fruit bodies of Amauroderma fungi comprise a cap and a stipe, and are typically woody, leathery, or corky in texture. The spores produced are usually spherical or nearly so, with a characteristic double wall structure that features U-shaped thickenings.
The fruit bodies of Amauroderma species are stipitate except in A. andina and may attain various shapes although centrally stipitate basidiocarps are most common. Several stipes may arise from the same base, frequently resulting in fused caps and compound fruit bodies. In section some fruit bodies are distinct with one or two distinct inner black bands or zones.
The stipe is often duplex with an outer dense layer surrounding an inner softer or hollow core sometimes separated by a black band. In species with a distinct tomentum on the stipe, there is often a dark zone just below the tomentum of the cap. These zones are absent from some species with a pale stipe without a tomentum. However, when present they continue into the context and frequently there is also another zone stretching more or less horizontally across the context.
Most basidiospores of Amauroderma mushrooms have an inner ornamented wall on which there is a hyaline (translucent) epicutis, which is very thin and difficult to see in ordinary microscopic preparations. Mature basidiospores are pale-yellowish. An apiculus
(a depressed area where the spore was once attached to the basidium via the sterigma) is often difficult to observe. Amauroderma is widespread in tropical areas.
Habitat loss from deforestation for oil palm plantation in tropical forests and partly collecting by local communities especially in Malaysia for medical properties. In Southeast Asia, more than 50% of Malaysia and Indonesia oil palm plantations in 2005 were on land that was forest in 1990 (Koh and Wilcove, 2008). Modification of substrates due to habitat loss will reduce population of species.
Amauroderma rugosum is a normally widespread in other regions but has been evaluated as a least concern (LC). This wild medicinal fungus always be consumed by indigenous communities in some countries.
Species can be found in tropical and subtropical areas. Collecting of this wild medicine fungus by human (Indigenous communities) and deforestation of diterocarp trees widely will reduce the population of fungus and degrade their natural habitat particulary in tropical rainforest regions in Thailand, Malaysia, Indonesia and Papua New Guinea
Population of species worldwide are facing habitat homogenization due to human activities especially in Asia. A low population density for this species in most regions due to extremely unfavorable habitats (substrate unfavourable for this saprophyte fungus) and habitat quality reduction then the species shows declines in some countries. Unclear of current trends of this species can be observed in most regions.
The species is growing as saprophytic (wood-decay fungi) on the ground with other plants and common in lowland dipterocarp of tropic forest and sub-tropic forest. The most suitable elevation for the growth and development of Amauroderma rugosum within on elevation of 200-1400m a.s.l. The higher humidity than 90% is the best for the growth and development of this species (Nguyen & Khanh, 2017).
In 2011 Conservation International listed the top 10 most endangered forests, characterized by having all lost 90% or more of their original habitat. Remaining habitat of endangered forests in Indo-Burma, New Caledonia, Sundaland, Philippines, Atlantic Forest, Mountains of Southwest China, California Floristic Province, Coastal Forests of Eastern Africa, Madagascar & Indian Ocean Islands and Eastern Afromontane only left between 11% and 5%.
Amauroderma rugosum is a wild mushroom that is worn as a necklace by the indigenous communities in Malaysia to prevent fits and incessant crying by babies.
Amauroderma rugosum, commonly known as “Jiǎzī” in China, is a wild mushroom traditionally used by the Chinese to reduce inflammation, to treat diuretic and upset stomach, and to prevent cancer.
It is also used by the indigenous communities in Malaysia to prevent epileptic episodes and incessant crying by babies. The aim of this study was to compare the wild and domesticated basidiocarps of A. rugosum for antioxidant and in vitro anti-inflammatory effects in LPS-stimulated RAW264.7 cells.
The wild basidiocarps of A. rugosum were collected from the Belum Forest, Perak, Malaysia and the domesticated basidiocarps of A. rugosum were cultivated in the mushroom house located in the University of Malaya, Kuala Lumpur, Malaysia. Both the wild and domesticated basidiocarps were subjected to ethanolic extraction and the extracts were tested for antioxidant and anti-inflammatory activities.
Amauroderma rugosum (AR) is a dietary mushroom in the Ganodermataceae family whose pharmacological activity and medicinal value have rarely been reported.
The antioxidant capacity and neuroprotective effects of AR were investigated. The aqueous extract of AR was confirmed to contain phenolic compounds, polysaccharides, and triterpenes. The results of 2,2-diphenyl-1-picryl-hydrazyl-hydrate (DPPH) and total antioxidant capacity assays revealed that AR extract scavenged reactive oxygen species. Moreover, AR extract decreased the cytotoxicity, oxidative stress, mitochondrial dysfunction, and apoptosis of PC12 cells induced by 6-hydroxydopamine (6-OHDA).
In addition, 6-OHDA upregulated the expressions of proapoptotic proteins and downregulated the Akt (protein kinase B)/mTOR- (mammalian target of rapamycin-) and MEK (mitogen-activated protein kinase kinase)/ERK- (extracellular signal-regulated kinases-) dependent signaling pathways. These effects of 6-OHDA were abolished or partially reversed by AR extract. Furthermore, the neuroprotective effects of AR in 6-OHDA-treated PC12 cells were significantly abolished by Akt and MEK inhibitor. Thus, AR extract possesses neuroprotective effects, probably through its antioxidant and antiapoptotic effects. These findings suggest the potential application of AR in the prevention or treatment of oxidative stress-related neurodegenerative diseases such as Parkinson’s disease.
To date, Parkinson’s disease remains the second most common neurodegenerative disease worldwide, and its incidence is increasing in people over 60 years of age. It is characterized by a selective and progressive loss of dopaminergic neurons in the substantia nigra pars compacta, leading to serious movement disturbances including postural instability, uncontrollable tremors, rigidity, and bradykinesia.
Currently, available treatments for Parkinson’s disease include dopaminergic replacement therapy and deep brain stimulation therapy. However, neither of these treatments can halt nor slow the progression of Parkinson’s disease. Therefore, it is crucial to develop novel drugs that can slow the neurodegenerative process.
Although the pathological mechanisms of Parkinson’s disease remain elusive, accumulating scientific evidence suggests that oxidative stress-induced cell injury plays an indispensable role in the degeneration of dopaminergic neurons. Interrupting the physiological maintenance of redox potential severely interferes with many biological processes in neurons, eventually leading to cell apoptosis.
Oxidative stress occurs when the rate of reactive oxygen species (ROS) scavenging is overwhelmed by the rate of ROS production. Excessive accumulation of ROS in dopaminergic neurons can damage most biological molecules, including lipids, proteins, and nucleic acids, thereby activating the intracellular inflammatory response, which induces cellular damage, mitochondrial dysfunction, oxidative DNA injury, and neuroinflammation. Therefore, decreasing oxidative injury in dopaminergic neurons has been widely proposed as an effective approach for the treatment of Parkinson’s disease.
Various experimental models have been established for investigating the role of oxidative stress in dopaminergic neuronal degeneration. These models involve the use of neurotoxins such as 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP), rotenone, 1,1-dimethyl-4,4-bipyridinium dichloride (paraquat) and 6-hydroxydopamine (6-OHDA).
The compound 6-OHDA, the hydroxylated analog of natural neurotransmitter dopamine, is a widely used neurotoxin that can be applied to establish different in vitro and in vivo Parkinson disease models. It is taken up by dopaminergic neurons via dopamine and norepinephrine transporters and is subsequently oxidized intracellularly, thereby releasing ROS including hydrogen peroxide, superoxide, and hydroxyl radicals.
Amauroderma rugosum (AR) is a basidiomycete in the Ganodermataceae family. This mushroom has a black stipe and a white surface covered with numerous pores. A notable characteristic of the mushroom is that its surface becomes red when it is scratched. Hence, it is also known as “blood Linzhi” in Chinese. Although AR is commonly consumed by people in China and South Asia, very few scientific studies have explored its beneficial effects on health or its medicinal value.
Nevertheless, a previous study reported that the extract of AR mycelia has antioxidant and anti-inflammatory effects in lipopolysaccharide-stimulated RAW 264.7 cells. Thus, AR might be a potential therapeutic agent or health supplement useful in the management of oxidative stress-related diseases. The aims of this study were to investigate the antioxidant capacity and neuroprotective activity of AR in a 6-OHDA-induced neurodegenerative cell model and to elucidate its underlying mechanisms of action.
Division: Basidiomycota
Class: Agaricomycetes
Order: Polyporales
Family: Ganodermataceae
Genus: Amauroderma
Murrill (1905)
Type species Amauroderma regulicolor
(Berk. ex Cooke) Murrill (1905)


Cyathus striatus
fluted bird's nest
fluted bird's nest

Cyathus striatus, commonly known as the fluted bird's nest, is a common saprobic bird's nest fungus with a widespread distribution throughout temperate regions of the world.
This fungus resembles a miniature bird's nest with numerous tiny "eggs"; the eggs, or peridioles, are actually lens-shaped bodies that contain spores.
C. striatus can be distinguished from most other bird's nest fungi by its hairy exterior and grooved (striated) inner walls. Although most frequently found growing on dead wood in open forests, it also grows on wood chip mulch in urban areas. The fruiting bodies are encountered from summer until early winter. The color and size of this species can vary somewhat, but they are typically less than a centimeter wide and tall, and grey or brown in color.
Another common name given to C. striatus, splash cups, alludes to the method of spore dispersal: the sides of the cup are angled such that falling drops of water can dislodge the peridioles and eject them from the cup. The specific epithet is derived from the Latin stria, meaning "with fine ridges or grooves".
The "nest", or peridium, is usually about 7 to 10 mm in height and 6 to 8 mm in width, but the size is somewhat variable and specimens have been found with heights and widths of up to 1.5 cm (0.59 in).
The shape typically resembles a vase or inverted cone. The outer surface (exoperidium) ranges in color from slightly brownish to grayish buff to deep brown; the exoperidium has a shaggy or hairy texture
(a tomentum), with the hairs mostly pointing downward. The inner surface of the peridium (the endoperidium) is striated or grooved, and shiny.
Young specimens have a lid, technically called an epiphragm, a thin membrane that covers the cup opening. The epiphragm is hairy like the rest of the exoperidial surface, but the hairs often wear off leaving behind a thin white layer stretched across the lid of the cup. As the peridium matures and expands, this membrane breaks and falls off, exposing the peridioles within. The peridium is attached to its growing surface by a mass of closely packed hyphae called an emplacement; in C. striatus the maximum diameter of the emplacement is typically 8–12 mm, and often incorporating small fragments of the growing surface into its structure. The species is inedible.
The peridioles structure are about 1 to 1.5 mm wide and rarely up to 2 mm wide. They are disc-shaped, but may appear angular due to pressure from neighbouring peridioles. Peridioles may be dark, or a drab gray if still covered with a thin membrane called a tunica.
Peridioles in C. striatus are sheathed and attached to the endoperidium by complex cords of mycelia known as a funiculus in the singular. The funiculus is differentiated into three regions: the basal piece, which attaches it to the inner wall of the peridium, the middle piece, and an upper sheath, called the purse, connected to the lower surface of the peridiole. Inside the purse and middle piece is a coiled thread of interwoven hyphae called the funicular cord, attached at one end to the peridiole and at the other end to an entangled mass of hyphae called the hapteron. When dry the funiculus is brittle, but when wet it is capable of long extension.
The basidia, the spore-bearing cells, are club-shaped with long stalks. They typically hold 4 spores that are sessile, that is, attached directly to the surface of the basidium, rather than by a short stalk (a sterigmata). Spores measure about 15 to 20 μm long by 8 to 12 μm wide. They are elliptical, smooth, hyaline, and notched at one end. During development, the spores are separated from the basidia when the latter collapse and gelatinize along with other cells lining the inner walls of the peridiole. The spores expand in size somewhat after being detached from the basidia.
Cyathus striatus is a saprobic fungus, deriving its nutrition from decaying organic material, and is typically found growing in clusters on small twigs or other woody debris. It is also common on mulch under shrubs. The features of the microenvironment largely influence the appearance of C. striatus; all else being equal, it is more likely to be found in moist, shallow depressions than elevated areas. It is very widespread in temperate areas throughout the world, growing in summer and fall. The fungus has been recorded from Asia, Europe, North America, Central America, South America, and New Zealand.
Cyathus striatus can reproduce both asexually (via vegetative spores), or sexually (with meiosis), typical of taxa in the basidiomycetes that contain both haploid and diploid stages.
Basidiospores produced in the peridioles each contain a single haploid nucleus. After the spores have been dispersed into a suitable growing environment, they germinate and develop into homokaryotic hyphae, with a single nucleus in each cell compartment. When two homokaryotic hyphae of different mating compatibility groups fuse with one another, they form a dikaryotic mycelia in a process called plasmogamy. After a period of time and under the appropriate environmental conditions, fruiting bodies may be formed from the dikaryotic mycelia. These fruiting bodies produce peridioles containing the basidia upon which new spores are made.
Young basidia contain a pair of haploid sexually compatible nuclei which fuse, and the resulting diploid fusion nucleus undergoes meiosis to produce haploid basidiospores. The process of meiosis in C. striatus has been found to be similar to that of higher organisms.
The cone shaped fruiting body of Cyathus striatus makes use of a splash-cup mechanism to help disperse the spores.
When a raindrop hits the interior of the cup with the optimal angle and velocity, the downward force of the water ejects the peridioles into the air. The force of ejection rips open the funiculus, releasing the tightly wound funicular cord. The hapteron attached to the end of the funiculus is adhesive, and when it contacts a nearby plant stem or stick, the hapteron sticks to it; the funicular cord wraps around the stem or stick powered by the force of the still-moving peridiole (similar to a tetherball). The peridioles degrade over time to eventually release the spores within, or they may be eaten by herbivorous animals and redeposited after passing through the digestive tract.
Cyathus striatus has proven to be a rich source of bioactive chemical compounds. It was first reported in 1971 to produce "indolic" substances (compounds with an indole ring structure) as well as a complex of diterpenoid antibiotic compounds collectively known as cyathins.
Several years later, research revealed the indolic substances to be compounds now known as striatins. Striatins (A, B and C) have antibiotic activity against fungi imperfecti, and various Gram-positive and Gram-negative bacteria.
C. striatus also produces sesquiterpene compounds called schizandronols. It also contains the triterpene compounds glochidone, glochidonol, glochidiol and glochidiol diacetate, cyathic acid, striatic acid, cyathadonic acid and epistriatic acid. The latter four compounds were unknown prior to their isolation from C. striatus.
Division: Basidiomycota
Class: Agaricomycetes
Order: Agaricales
Family: Nidulariaceae
Genus: Cyathus
Species: C. striatus
Binomial name Cyathus striatus
(Huds.) Willd. (1787)
This fungus resembles a miniature bird's nest with numerous tiny "eggs"; the eggs, or peridioles, are actually lens-shaped bodies that contain spores.
C. striatus can be distinguished from most other bird's nest fungi by its hairy exterior and grooved (striated) inner walls. Although most frequently found growing on dead wood in open forests, it also grows on wood chip mulch in urban areas. The fruiting bodies are encountered from summer until early winter. The color and size of this species can vary somewhat, but they are typically less than a centimeter wide and tall, and grey or brown in color.
Another common name given to C. striatus, splash cups, alludes to the method of spore dispersal: the sides of the cup are angled such that falling drops of water can dislodge the peridioles and eject them from the cup. The specific epithet is derived from the Latin stria, meaning "with fine ridges or grooves".
The "nest", or peridium, is usually about 7 to 10 mm in height and 6 to 8 mm in width, but the size is somewhat variable and specimens have been found with heights and widths of up to 1.5 cm (0.59 in).
The shape typically resembles a vase or inverted cone. The outer surface (exoperidium) ranges in color from slightly brownish to grayish buff to deep brown; the exoperidium has a shaggy or hairy texture
(a tomentum), with the hairs mostly pointing downward. The inner surface of the peridium (the endoperidium) is striated or grooved, and shiny.
Young specimens have a lid, technically called an epiphragm, a thin membrane that covers the cup opening. The epiphragm is hairy like the rest of the exoperidial surface, but the hairs often wear off leaving behind a thin white layer stretched across the lid of the cup. As the peridium matures and expands, this membrane breaks and falls off, exposing the peridioles within. The peridium is attached to its growing surface by a mass of closely packed hyphae called an emplacement; in C. striatus the maximum diameter of the emplacement is typically 8–12 mm, and often incorporating small fragments of the growing surface into its structure. The species is inedible.
The peridioles structure are about 1 to 1.5 mm wide and rarely up to 2 mm wide. They are disc-shaped, but may appear angular due to pressure from neighbouring peridioles. Peridioles may be dark, or a drab gray if still covered with a thin membrane called a tunica.
Peridioles in C. striatus are sheathed and attached to the endoperidium by complex cords of mycelia known as a funiculus in the singular. The funiculus is differentiated into three regions: the basal piece, which attaches it to the inner wall of the peridium, the middle piece, and an upper sheath, called the purse, connected to the lower surface of the peridiole. Inside the purse and middle piece is a coiled thread of interwoven hyphae called the funicular cord, attached at one end to the peridiole and at the other end to an entangled mass of hyphae called the hapteron. When dry the funiculus is brittle, but when wet it is capable of long extension.
The basidia, the spore-bearing cells, are club-shaped with long stalks. They typically hold 4 spores that are sessile, that is, attached directly to the surface of the basidium, rather than by a short stalk (a sterigmata). Spores measure about 15 to 20 μm long by 8 to 12 μm wide. They are elliptical, smooth, hyaline, and notched at one end. During development, the spores are separated from the basidia when the latter collapse and gelatinize along with other cells lining the inner walls of the peridiole. The spores expand in size somewhat after being detached from the basidia.
Cyathus striatus is a saprobic fungus, deriving its nutrition from decaying organic material, and is typically found growing in clusters on small twigs or other woody debris. It is also common on mulch under shrubs. The features of the microenvironment largely influence the appearance of C. striatus; all else being equal, it is more likely to be found in moist, shallow depressions than elevated areas. It is very widespread in temperate areas throughout the world, growing in summer and fall. The fungus has been recorded from Asia, Europe, North America, Central America, South America, and New Zealand.
Cyathus striatus can reproduce both asexually (via vegetative spores), or sexually (with meiosis), typical of taxa in the basidiomycetes that contain both haploid and diploid stages.
Basidiospores produced in the peridioles each contain a single haploid nucleus. After the spores have been dispersed into a suitable growing environment, they germinate and develop into homokaryotic hyphae, with a single nucleus in each cell compartment. When two homokaryotic hyphae of different mating compatibility groups fuse with one another, they form a dikaryotic mycelia in a process called plasmogamy. After a period of time and under the appropriate environmental conditions, fruiting bodies may be formed from the dikaryotic mycelia. These fruiting bodies produce peridioles containing the basidia upon which new spores are made.
Young basidia contain a pair of haploid sexually compatible nuclei which fuse, and the resulting diploid fusion nucleus undergoes meiosis to produce haploid basidiospores. The process of meiosis in C. striatus has been found to be similar to that of higher organisms.
The cone shaped fruiting body of Cyathus striatus makes use of a splash-cup mechanism to help disperse the spores.
When a raindrop hits the interior of the cup with the optimal angle and velocity, the downward force of the water ejects the peridioles into the air. The force of ejection rips open the funiculus, releasing the tightly wound funicular cord. The hapteron attached to the end of the funiculus is adhesive, and when it contacts a nearby plant stem or stick, the hapteron sticks to it; the funicular cord wraps around the stem or stick powered by the force of the still-moving peridiole (similar to a tetherball). The peridioles degrade over time to eventually release the spores within, or they may be eaten by herbivorous animals and redeposited after passing through the digestive tract.
Cyathus striatus has proven to be a rich source of bioactive chemical compounds. It was first reported in 1971 to produce "indolic" substances (compounds with an indole ring structure) as well as a complex of diterpenoid antibiotic compounds collectively known as cyathins.
Several years later, research revealed the indolic substances to be compounds now known as striatins. Striatins (A, B and C) have antibiotic activity against fungi imperfecti, and various Gram-positive and Gram-negative bacteria.
C. striatus also produces sesquiterpene compounds called schizandronols. It also contains the triterpene compounds glochidone, glochidonol, glochidiol and glochidiol diacetate, cyathic acid, striatic acid, cyathadonic acid and epistriatic acid. The latter four compounds were unknown prior to their isolation from C. striatus.
Division: Basidiomycota
Class: Agaricomycetes
Order: Agaricales
Family: Nidulariaceae
Genus: Cyathus
Species: C. striatus
Binomial name Cyathus striatus
(Huds.) Willd. (1787)








Tulostoma exasperatum var. ridleyi
Stalkballs or Stalked puffballs
Stalkballs or Stalked puffballs

Tulostoma is a genus of over 100 species of fungi in the family Agaricaceae. Commonly known as
stalkballs or stalked puffballs, the cosmopolitan genus consists of species which produce small fruit bodies, characterized by stalks inserted in a socket at the base of the spherical spore-sac opened by a small and apical mouth.
The spore-sac contains gleba, a mixture of spores and associated cells; at maturity, the spores are released through one or more apical pores.
Tulostoma species prefer xeric microhabitats, savannahs and deserts, and are saprobic—obtaining nutrients by decomposing roots, buried wood and other organic material of plant origin.
Tulostoma exasperatum is pantropical while var. ridleyi is endemic to South East Asia.
According to Wright (1987), Tulostoma exasperatum var. ridleyi (Massee) J.E. Wright has different features from the norm: the stems are longer, reaching more than 120 mm in length, and the basidoma is more robust.
The spores are also larger 4.7-5.1 x 6.4-8.1 microns. This variety is supposedly restricted to S.E. Asia. Further study may be needed to determine if this is true.
Kingdom: Fungi
Phylum: Basidiomycota
Class: Agaricomycetes
Order: Agaricales
Family: Agaricaceae
Common names: Stalkballs or Stalked puffballs.
Scientific name: Tulostoma exasperatum var. ridleyi
Author: (Massee) J.E. Wright
IUCN Specialist Group: Mushroom, Bracket and Puffball
stalkballs or stalked puffballs, the cosmopolitan genus consists of species which produce small fruit bodies, characterized by stalks inserted in a socket at the base of the spherical spore-sac opened by a small and apical mouth.
The spore-sac contains gleba, a mixture of spores and associated cells; at maturity, the spores are released through one or more apical pores.
Tulostoma species prefer xeric microhabitats, savannahs and deserts, and are saprobic—obtaining nutrients by decomposing roots, buried wood and other organic material of plant origin.
Tulostoma exasperatum is pantropical while var. ridleyi is endemic to South East Asia.
According to Wright (1987), Tulostoma exasperatum var. ridleyi (Massee) J.E. Wright has different features from the norm: the stems are longer, reaching more than 120 mm in length, and the basidoma is more robust.
The spores are also larger 4.7-5.1 x 6.4-8.1 microns. This variety is supposedly restricted to S.E. Asia. Further study may be needed to determine if this is true.
Kingdom: Fungi
Phylum: Basidiomycota
Class: Agaricomycetes
Order: Agaricales
Family: Agaricaceae
Common names: Stalkballs or Stalked puffballs.
Scientific name: Tulostoma exasperatum var. ridleyi
Author: (Massee) J.E. Wright
IUCN Specialist Group: Mushroom, Bracket and Puffball

Filoboletus manipularis
Bioluminescence glow mushroom
Bioluminescence glow mushroom

Mycena is a large genus of small saprotrophic mushrooms that are rarely more than a few centimeters in width. They are characterized by a white spore print.
Mycena manipularis is a species of agaric fungus in the family Mycenaceae. Found in Australasia, Malaysia, Singapore and the Pacific islands, the mycelium and fruit bodies of the fungus are bioluminescent.
Filoboletus manipularis is a luminescent fungal species widely distributed on rotting wood throughout Asian, Australasian and Pacific tropical regions.
The phylogenetic relationships among species accommodated in Filoboletus have not been studied, and even their generic affinities remain largely unresolved.
Mycobank gives Favolaschia as the currently accepted generic placement for the species. However, phylogenetic analysis of available sequences place F. manipularis within a Mycena clade closest to M. rubreomarginata and distant from Favolaschia species.
According to Desjardins et al. (2008), F. manipularis and related species currently assigned to Filoboletus and Poromycena require a new generic name since none are closely related to the type species of these genera. the current study was undertaken to determine if the morphological variation observed in F. manipularis represents more than one species or could be correlated to the infraspecific phylogenetics of the species.
Filoboletus manipularis is easily identified by its white to pale cream cap with rounded pores (not gills) on its under surface. Caps are 5-35 mm diam, with most around 20 mm diam., convex to bell-shaped becoming umbonate (with central, raised knob), and often appear dimpled due to the pores. The centre of the cap often has a brown tint.Gills: Has rounded pores (not gills). Pores are adnate, 1-3 per mm and symmetrically arranged. The central stem is up to 80 mm high and 2-5 mm diam, white but pale brown towards the slightly swollen base and it has a faint glow.
F. manipularis emits a faint white luminance when see in total darkness and one will by pass it unknowingly that this species are bioluminescent, compare to N.nambi its faint white luminance is slightly brighter than F. manipularis and M.chlorophos is the species that emits the strongest green luminance in darkness and one will definately see it even from a distance.
F.manipularis grows singly but usually in clusters (caespitose). It occurs in subtropical to tropical rainforests. The morphological variation of basidiomata of Filoboletus manipularis (Berk) Singer collected in southern Viet Nam was studied.
Phylogenetic analyses comprising three gene loci indicated that these collections, although exhibiting widely varying morphologies, represented a single species with a population composed of genetically diverse, sexually compatible monokaryon parental strains. No correlation was found between any aspect of morphological variation and intraspecific phylogenetic patterns for the three gene loci studied.
Primers were designed to amplify the intron-rich 5′ region of the translation elongation factor 1-α gene (tef1α) and amplicons cloned and sequenced to characterize the parental haplotypes for individual basidiomata. The presence of recombination over the entire morphological diversity seen was confirmed by split decomposition analysis and analysis of gene diversity indicated a lack of allelic fixation within local populations. On several occasions, more than two apparent parental haplotypes were characterized from individual basidiomata, indicating that at least some basidiomata are chimeric or otherwise develop from a multinucleate condition. The literature supporting observations of the occurrence of multinucleate basidiomata is reviewed and possible mechanisms for this phenomenon are proposed.
Division: Basidiomycota
Class: Agaricomycetes
Order: Agaricales
Family: Mycenaceae
Genus: Mycena
Species: M. manipularis
Binomial name Mycena manipularis
(Berk.) Sacc. (1887)
Mycena manipularis is a species of agaric fungus in the family Mycenaceae. Found in Australasia, Malaysia, Singapore and the Pacific islands, the mycelium and fruit bodies of the fungus are bioluminescent.
Filoboletus manipularis is a luminescent fungal species widely distributed on rotting wood throughout Asian, Australasian and Pacific tropical regions.
The phylogenetic relationships among species accommodated in Filoboletus have not been studied, and even their generic affinities remain largely unresolved.
Mycobank gives Favolaschia as the currently accepted generic placement for the species. However, phylogenetic analysis of available sequences place F. manipularis within a Mycena clade closest to M. rubreomarginata and distant from Favolaschia species.
According to Desjardins et al. (2008), F. manipularis and related species currently assigned to Filoboletus and Poromycena require a new generic name since none are closely related to the type species of these genera. the current study was undertaken to determine if the morphological variation observed in F. manipularis represents more than one species or could be correlated to the infraspecific phylogenetics of the species.
Filoboletus manipularis is easily identified by its white to pale cream cap with rounded pores (not gills) on its under surface. Caps are 5-35 mm diam, with most around 20 mm diam., convex to bell-shaped becoming umbonate (with central, raised knob), and often appear dimpled due to the pores. The centre of the cap often has a brown tint.Gills: Has rounded pores (not gills). Pores are adnate, 1-3 per mm and symmetrically arranged. The central stem is up to 80 mm high and 2-5 mm diam, white but pale brown towards the slightly swollen base and it has a faint glow.
F. manipularis emits a faint white luminance when see in total darkness and one will by pass it unknowingly that this species are bioluminescent, compare to N.nambi its faint white luminance is slightly brighter than F. manipularis and M.chlorophos is the species that emits the strongest green luminance in darkness and one will definately see it even from a distance.
F.manipularis grows singly but usually in clusters (caespitose). It occurs in subtropical to tropical rainforests. The morphological variation of basidiomata of Filoboletus manipularis (Berk) Singer collected in southern Viet Nam was studied.
Phylogenetic analyses comprising three gene loci indicated that these collections, although exhibiting widely varying morphologies, represented a single species with a population composed of genetically diverse, sexually compatible monokaryon parental strains. No correlation was found between any aspect of morphological variation and intraspecific phylogenetic patterns for the three gene loci studied.
Primers were designed to amplify the intron-rich 5′ region of the translation elongation factor 1-α gene (tef1α) and amplicons cloned and sequenced to characterize the parental haplotypes for individual basidiomata. The presence of recombination over the entire morphological diversity seen was confirmed by split decomposition analysis and analysis of gene diversity indicated a lack of allelic fixation within local populations. On several occasions, more than two apparent parental haplotypes were characterized from individual basidiomata, indicating that at least some basidiomata are chimeric or otherwise develop from a multinucleate condition. The literature supporting observations of the occurrence of multinucleate basidiomata is reviewed and possible mechanisms for this phenomenon are proposed.
Division: Basidiomycota
Class: Agaricomycetes
Order: Agaricales
Family: Mycenaceae
Genus: Mycena
Species: M. manipularis
Binomial name Mycena manipularis
(Berk.) Sacc. (1887)
How is bioluminescence form
Bioluminescence is the production and emission of light by a living organism. It is a form of chemiluminescence.
Bioluminescence occurs widely in marine vertebrates and invertebrates, as well as in some fungi, microorganisms including some bioluminescent bacteria, and terrestrial arthropods such as fireflies.
Bioluminescence is used by living things to hunt prey, defend against predators, find mates, and execute other vital activities and some species luminesce to confuse attackers.
Most bioluminescent organisms are found in the ocean. These bioluminescent marine species include fish, bacteria, and jellies and Some bioluminescent organisms, including fireflies and fungi, are found on land. There are almost no bioluminescent organisms native to freshwater habitats.
In some animals, the light is bacteriogenic, produced by symbiotic bacteria such as those from the genus Vibrio; in others, it is autogenic, produced by the animals themselves.
Bioluminescence is a "cold light." Cold light means less than 20% of the light generates thermal radiation, or heat.
Luciferin is the compound that actually produces light. Luciferase is an enzyme. An enzyme is a chemical (called a catalyst) that interacts with a substrate to affect the rate of a chemical reaction.
In a general sense, the principal chemical reaction in bioluminescence involves a light-emitting molecule and an enzyme, generally called luciferin and luciferase, respectively.
In a chemical reaction, luciferin is called the substrate. The bioluminescent color (yellow in fireflies, greenish in lanternfish) is a result of the arrangement of luciferin molecules.
Some bioluminescent organisms produce (synthesize) luciferin on their own and some bioluminescent organisms do not synthesize luciferin. Instead, they absorb it through other organisms, either as food or in a symbiotic relationship.
The interaction of the luciferase with oxidized (oxygen-added) luciferin creates a byproduct, called oxyluciferin which is a type of chemical reaction creates light.
Most bioluminescent reactions involve luciferin and luciferase. Some reactions, however, do not involve an enzyme (luciferase). These reactions involve a chemical called a photoprotein. Photoproteins combine with luciferins and oxygen, but need another agent, often an ion of the element calcium, to produce light.
The appearance of bioluminescent light varies greatly, depending on the habitat and organism in which it is found.
Some species of fungi present in decaying wood, for instance, emit a fairly consistent glow, called foxfire and some organisms emit very bright light continuously.
Bioluminescence occurs widely in marine vertebrates and invertebrates, as well as in some fungi, microorganisms including some bioluminescent bacteria, and terrestrial arthropods such as fireflies.
Bioluminescence is used by living things to hunt prey, defend against predators, find mates, and execute other vital activities and some species luminesce to confuse attackers.
Most bioluminescent organisms are found in the ocean. These bioluminescent marine species include fish, bacteria, and jellies and Some bioluminescent organisms, including fireflies and fungi, are found on land. There are almost no bioluminescent organisms native to freshwater habitats.
In some animals, the light is bacteriogenic, produced by symbiotic bacteria such as those from the genus Vibrio; in others, it is autogenic, produced by the animals themselves.
Bioluminescence is a "cold light." Cold light means less than 20% of the light generates thermal radiation, or heat.
Luciferin is the compound that actually produces light. Luciferase is an enzyme. An enzyme is a chemical (called a catalyst) that interacts with a substrate to affect the rate of a chemical reaction.
In a general sense, the principal chemical reaction in bioluminescence involves a light-emitting molecule and an enzyme, generally called luciferin and luciferase, respectively.
In a chemical reaction, luciferin is called the substrate. The bioluminescent color (yellow in fireflies, greenish in lanternfish) is a result of the arrangement of luciferin molecules.
Some bioluminescent organisms produce (synthesize) luciferin on their own and some bioluminescent organisms do not synthesize luciferin. Instead, they absorb it through other organisms, either as food or in a symbiotic relationship.
The interaction of the luciferase with oxidized (oxygen-added) luciferin creates a byproduct, called oxyluciferin which is a type of chemical reaction creates light.
Most bioluminescent reactions involve luciferin and luciferase. Some reactions, however, do not involve an enzyme (luciferase). These reactions involve a chemical called a photoprotein. Photoproteins combine with luciferins and oxygen, but need another agent, often an ion of the element calcium, to produce light.
The appearance of bioluminescent light varies greatly, depending on the habitat and organism in which it is found.
Some species of fungi present in decaying wood, for instance, emit a fairly consistent glow, called foxfire and some organisms emit very bright light continuously.
Why Do Some Mushrooms Glow In The Dark?
Why Do Some Mushrooms Glow In The Dark?
Glowing in the dark costs energy.
The Secret Behind Bioluminescent Mushrooms’ Magic Glow.
Scientists use chemistry to account for an astonishing phenomenon. Researchers described the compound that gives the mushrooms their glow — It’s called oxyluciferin.
Mushrooms use luciferins—light-emitting compounds found in other glowing animals and plants—to attract insects. The bugs then help spread their spores to sheltered places in the forest, which helps the mushroom species survive.
Luciferins give fireflies and even bioluminescent underwater creatures their glow. Paired with an enzyme and oxygen, it releases light that illuminates the fungi.
Scientists went foraging for the glow-in-the-dark mushrooms and they crushed the mushrooms to make a slurry filled with luciferins. Then they isolated the luciferin and studied it, capturing its chemical structure and experimenting with its ability to fuel those flourescent colors.
Not only does the team now know that the mushrooms are fueled by their own kind of luciferin, but they also figured out that the enzyme that combines with the chemical to trigger light could be what they call “promiscuous.”
That means that the enzyme might be able to interact with different luciferins—and produce even more shades of that pretty glow. And that suggests that when it comes to these magical mushrooms, there’s even more to discover.
Fungi produce tiny spores to spread themselves, much like seeds from trees. Insects can help transport the fungal spores. Fruit bodies are produced to disperse their sexual progeny as spores. Many fungi shoot spores into the air from the underside of the mushrooms, relying on moving air currents to passively distribute the spores over a wide area.
Through aroma. Truffles, the fruiting body of the Ascomycete fungi, use their smell to attract fungivores such as pigs or squirrels who eat them and leave spores behind in their waste. Stinkhorn mushrooms have a foul-smelling slime which attracts flies and other insects. The flies eat the slime and unwittingly spread the spores elsewhere.
Since bioluminescence is so energetically costly, the mushrooms only glow at night. They don’t glow bright enough during the day to be extra visible. An added bonus is that spores prefer to become active and grow at night when it is more humid.
Bioluminescence is one of the factors attracting insects to the mushrooms to help them spread their spores.
"On a totally dark night, without any moon, if you have your light off, these green mushrooms are basically the only light source you see in the forest besides the fireflies."
You just have to turn off your flashlight and the mushrooms stand out if they're there.
Glowing in the dark costs energy.
The Secret Behind Bioluminescent Mushrooms’ Magic Glow.
Scientists use chemistry to account for an astonishing phenomenon. Researchers described the compound that gives the mushrooms their glow — It’s called oxyluciferin.
Mushrooms use luciferins—light-emitting compounds found in other glowing animals and plants—to attract insects. The bugs then help spread their spores to sheltered places in the forest, which helps the mushroom species survive.
Luciferins give fireflies and even bioluminescent underwater creatures their glow. Paired with an enzyme and oxygen, it releases light that illuminates the fungi.
Scientists went foraging for the glow-in-the-dark mushrooms and they crushed the mushrooms to make a slurry filled with luciferins. Then they isolated the luciferin and studied it, capturing its chemical structure and experimenting with its ability to fuel those flourescent colors.
Not only does the team now know that the mushrooms are fueled by their own kind of luciferin, but they also figured out that the enzyme that combines with the chemical to trigger light could be what they call “promiscuous.”
That means that the enzyme might be able to interact with different luciferins—and produce even more shades of that pretty glow. And that suggests that when it comes to these magical mushrooms, there’s even more to discover.
Fungi produce tiny spores to spread themselves, much like seeds from trees. Insects can help transport the fungal spores. Fruit bodies are produced to disperse their sexual progeny as spores. Many fungi shoot spores into the air from the underside of the mushrooms, relying on moving air currents to passively distribute the spores over a wide area.
Through aroma. Truffles, the fruiting body of the Ascomycete fungi, use their smell to attract fungivores such as pigs or squirrels who eat them and leave spores behind in their waste. Stinkhorn mushrooms have a foul-smelling slime which attracts flies and other insects. The flies eat the slime and unwittingly spread the spores elsewhere.
Since bioluminescence is so energetically costly, the mushrooms only glow at night. They don’t glow bright enough during the day to be extra visible. An added bonus is that spores prefer to become active and grow at night when it is more humid.
Bioluminescence is one of the factors attracting insects to the mushrooms to help them spread their spores.
"On a totally dark night, without any moon, if you have your light off, these green mushrooms are basically the only light source you see in the forest besides the fireflies."
You just have to turn off your flashlight and the mushrooms stand out if they're there.

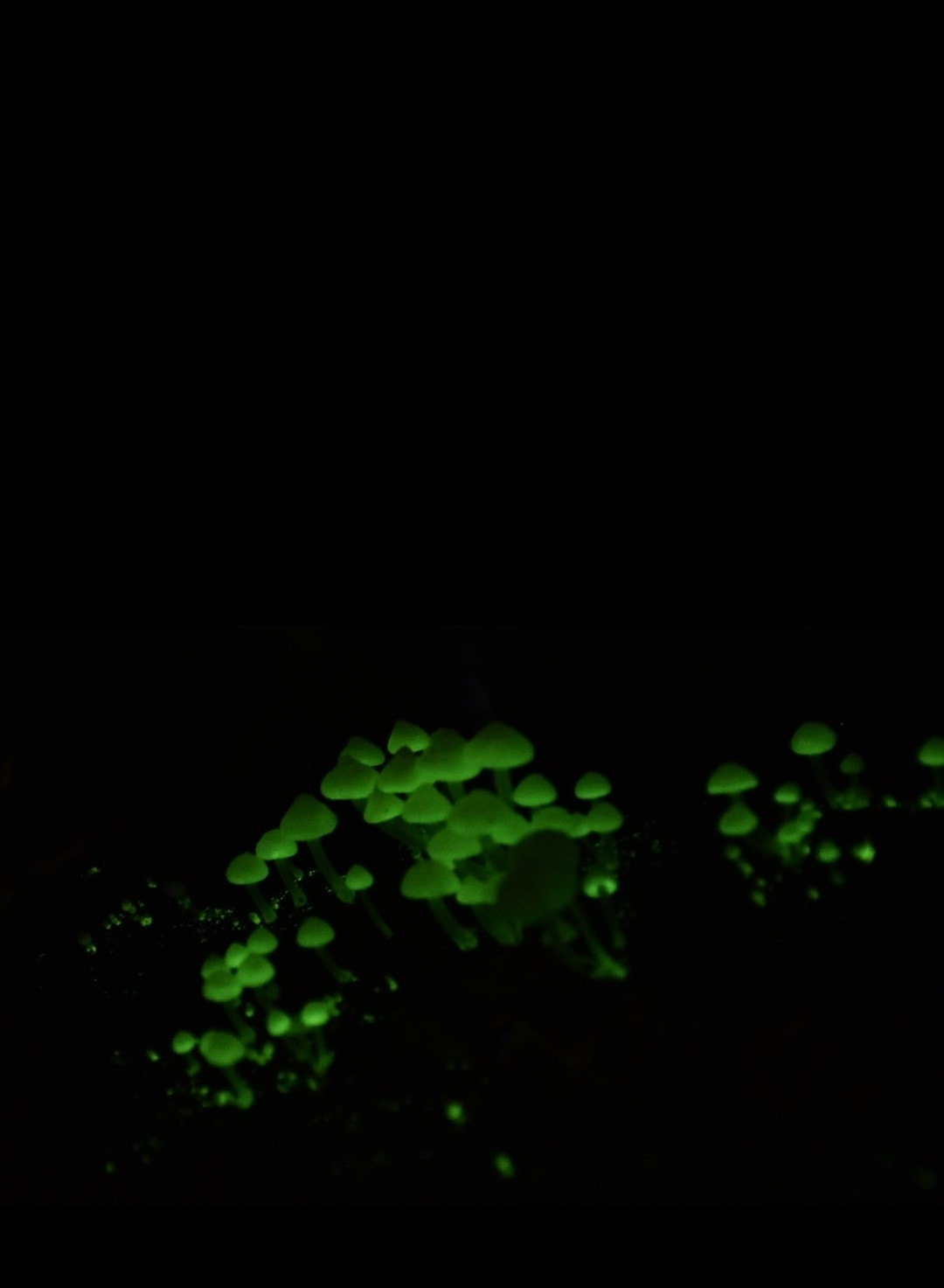

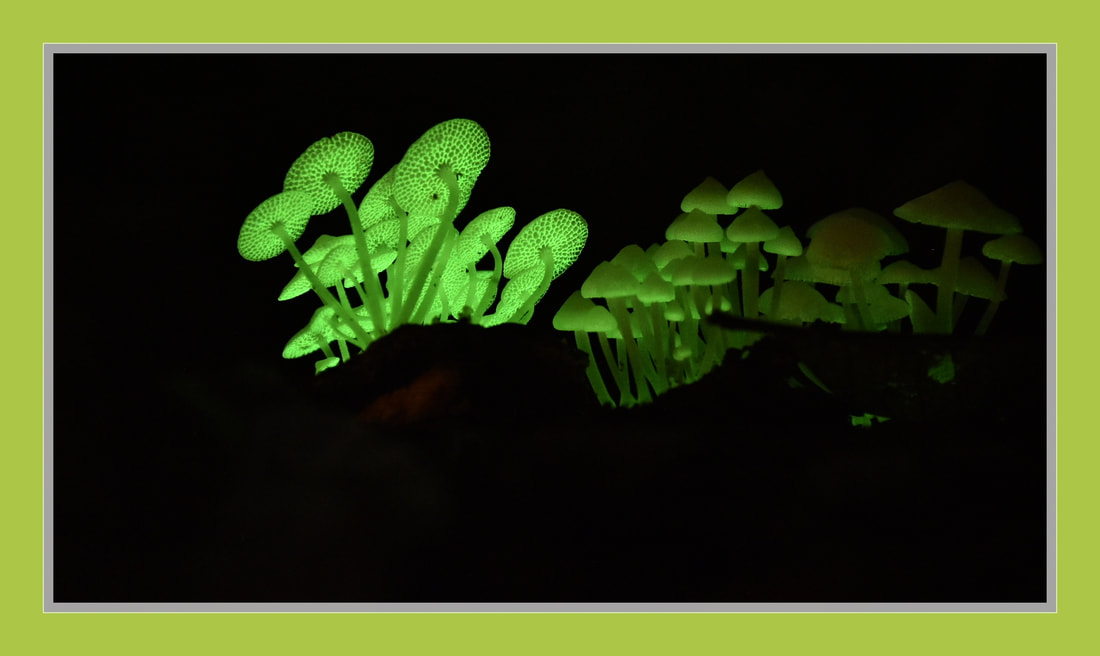
Marasmius siccus
orange pinwheel
orange pinwheel

Marasmius siccus, or orange pinwheel, is a small orange mushroom in the Marasmius genus, with a "beach umbrella"-shaped cap. The tough shiny bare stem is pale at the top but reddish brown below, and the gills are whitish. The stem is 3–7 centimetres (1.2–2.8 in) tall and the cap is 0.5–2.5 centimetres (0.20–0.98 in) wide.
At a microscopic level, the club-shaped spores are very long and thin, being roughly 19 µm by 4 µm. The distinctive cheilocystidia are broadly club-shaped with finger-like protrusions at the far end. Such cells also sometimes occur in other related mushrooms and they are known as "broom cells of the siccus type".
This mushroom is found in hardwood forests from the Rocky Mountains to the Appalachian Mountains and also in northern Europe and Asia.
Although nonpoisonous, they are too small to be considered worthwhile as food.
Kingdom: Fungi
Division: Basidiomycota
Class: Agaricomycete
Order: Agaricales
Family: Marasmiaceae
Genus: Marasmius
Species: M. siccus
Binomial name Marasmius siccus
Schwein. ex Fr.
At a microscopic level, the club-shaped spores are very long and thin, being roughly 19 µm by 4 µm. The distinctive cheilocystidia are broadly club-shaped with finger-like protrusions at the far end. Such cells also sometimes occur in other related mushrooms and they are known as "broom cells of the siccus type".
This mushroom is found in hardwood forests from the Rocky Mountains to the Appalachian Mountains and also in northern Europe and Asia.
Although nonpoisonous, they are too small to be considered worthwhile as food.
Kingdom: Fungi
Division: Basidiomycota
Class: Agaricomycete
Order: Agaricales
Family: Marasmiaceae
Genus: Marasmius
Species: M. siccus
Binomial name Marasmius siccus
Schwein. ex Fr.


The Pezizaceae
Cup fungi
Cup fungi

The Pezizaceae (commonly referred to as cup fungi) are a family of fungi in the Ascomycota which produce mushrooms that tends to grow in the shape of a "cup". Spores are formed on the inner surface of the fruit body (mushroom).
The cup shape typically serves to focus raindrops into splashing spores out of the cup. Additionally, the curvature enables wind currents to blow the spores out in a different manner than in most agarics and boletes.
Cup fungi grow in peculiar shapes, frequently resembling cups or saucers. For example, the orange peel fungus (Aleuria aurantia) resembles a discarded orange rind. They may be vividly colored, like the scarlet cup (Sarcoscypha coccinea), which is often one of the first signs of spring where it grows. According to one 2008 estimate, the family contains 31 genera and 230 species.
Cookeina is a genus of cup fungi in the family Sarcoscyphaceae, members of which may be found in tropical and subtropical regions of the world. Species may be found on fallen branches of angiosperms, trunks, and sometimes on fruits.
The Temuans of Peninsular Malaysia are reported to use certain species from this genus as food, and also as a bait for fishing, where it is rubbed against the hook.
Species in the Cookeina have a deep, cup-shaped to funnel-shaped fruiting bodies, or apothecia. The inner spore-bearing surface of the apothecium, the hymenium, is brightly colored, yellow to red, although the color will fade upon drying. The outer surface is less brightly colored. The excipulum, the tissue making up the walls of the apothecium, is thin and flexible. When hairs are present on the apothecium, they are fasciculate―made of bundles of cylindrical hyphae.
The Cookeina have asci which are constricted abruptly below and form a blunt, rounded base with a slim, tail-like connection. They have asci which mature simultaneously rather than in series. They have paraphyses which anastomose and form a three-dimensional network. Ascospores are large (20—40 µm long), ellipsoidal or slightly unequal-sided, and either smooth or ornamented with fine wrinkles.
The genus appears to be restricted to wood, commonly to wood in early stages of decay.
When mature apothecia become filled with water, the asci absorb some of that water and develop a Turgor pressure, a hydrostatic pressure within the ascus which put pressure on the tip of the ascus, held in place by the rigid ascus wall. As the water level in the cup reduces due to evaporation, the asci tips dry out, resulting in a negative vapor pressure that ultimately results in the thin tissue at the wall of the apex (the operculum) breaking outward, releasing the spores.
Phylogenetic analyses of ribosomal DNA has helped to clarify the evolutionary and genetic relationships amongst the species in Cookeina. The species C. speciosa, C. tricholoma, and C. sinensis belong to a monophyletic group, and all are hairy, with stipes, and lack a well-defined layer of slime in the excipulum at maturity.
Another monophyletic grouping contains the species C. Venezuela and C. colensoi, which do have a slime layer on the excipulum, and have either short stipes or not at all (sessile). In this analysis, C. indica and C. insititia did not clearly resolve with the other clades.
Kingdom: Fungi
Division: Ascomycota
Class: Pezizomycetes
Subclass: Pezizomycetidae
Order: Pezizales
Family: Sarcoscyphaceae
Genus: Cookeina
Kuntze (1891)
The cup shape typically serves to focus raindrops into splashing spores out of the cup. Additionally, the curvature enables wind currents to blow the spores out in a different manner than in most agarics and boletes.
Cup fungi grow in peculiar shapes, frequently resembling cups or saucers. For example, the orange peel fungus (Aleuria aurantia) resembles a discarded orange rind. They may be vividly colored, like the scarlet cup (Sarcoscypha coccinea), which is often one of the first signs of spring where it grows. According to one 2008 estimate, the family contains 31 genera and 230 species.
Cookeina is a genus of cup fungi in the family Sarcoscyphaceae, members of which may be found in tropical and subtropical regions of the world. Species may be found on fallen branches of angiosperms, trunks, and sometimes on fruits.
The Temuans of Peninsular Malaysia are reported to use certain species from this genus as food, and also as a bait for fishing, where it is rubbed against the hook.
Species in the Cookeina have a deep, cup-shaped to funnel-shaped fruiting bodies, or apothecia. The inner spore-bearing surface of the apothecium, the hymenium, is brightly colored, yellow to red, although the color will fade upon drying. The outer surface is less brightly colored. The excipulum, the tissue making up the walls of the apothecium, is thin and flexible. When hairs are present on the apothecium, they are fasciculate―made of bundles of cylindrical hyphae.
The Cookeina have asci which are constricted abruptly below and form a blunt, rounded base with a slim, tail-like connection. They have asci which mature simultaneously rather than in series. They have paraphyses which anastomose and form a three-dimensional network. Ascospores are large (20—40 µm long), ellipsoidal or slightly unequal-sided, and either smooth or ornamented with fine wrinkles.
The genus appears to be restricted to wood, commonly to wood in early stages of decay.
When mature apothecia become filled with water, the asci absorb some of that water and develop a Turgor pressure, a hydrostatic pressure within the ascus which put pressure on the tip of the ascus, held in place by the rigid ascus wall. As the water level in the cup reduces due to evaporation, the asci tips dry out, resulting in a negative vapor pressure that ultimately results in the thin tissue at the wall of the apex (the operculum) breaking outward, releasing the spores.
Phylogenetic analyses of ribosomal DNA has helped to clarify the evolutionary and genetic relationships amongst the species in Cookeina. The species C. speciosa, C. tricholoma, and C. sinensis belong to a monophyletic group, and all are hairy, with stipes, and lack a well-defined layer of slime in the excipulum at maturity.
Another monophyletic grouping contains the species C. Venezuela and C. colensoi, which do have a slime layer on the excipulum, and have either short stipes or not at all (sessile). In this analysis, C. indica and C. insititia did not clearly resolve with the other clades.
Kingdom: Fungi
Division: Ascomycota
Class: Pezizomycetes
Subclass: Pezizomycetidae
Order: Pezizales
Family: Sarcoscyphaceae
Genus: Cookeina
Kuntze (1891)




Ramaria stricta
Strict-branch coral fungus
Strict-branch coral fungus

Ramaria stricta, commonly known as the strict-branch coral is a coral fungus of the genus Ramaria. It has a cosmopolitan distribution, and grows on dead wood, stumps, trunks, and branches of both deciduous and coniferous trees.
Its fruit body is up to 10 cm (3.9 in) tall, made of multiple slender, compact, and vertical parallel branches. Its color is typically light tan to vinaceous-brown. All parts of the mushroom will bruise when handled.
There are several lookalike corals that can usually be distinguished from
R. stricta by differences in coloration, bruising reaction, or microscopic features. The fungus is inedible due to its unpleasant odor and bitter taste.
The species was originally described under the name Clavaria stricta by Christian Hendrik Persoon in 1795. In 1888, French mycologist Lucien Quélet transferred the species to the genus Ramaria. It is commonly known as the "upright coral".
The color of the fruit body is light yellow brown to paler toward extremities, the tips are light yellow; bruising light reddish brown. The basidiocarp has a leathery texture when fresh, but becomes brittle when dry. The stipe is branched up to 8 times, and the branches are all upright and nearly parallel. The branches ending in 4 to 5 thornlike tips. Overall, the fruit body appears bushy, and is medium-sized, up to 10 by 7 cm (3.9 by 2.8 in). The stipe is single or branching from the base; with white mycelium and rhizomorphs radiating from the base. The odor is of anise. The taste is bitter. The fruit bodies have been described as "edible but unpalatable," although other guides say the species is inedible.
The spore print is dark yellow. Spores are roughly elliptical, dotted with low cyanophilous warts, and measure 7–10 by 3.5–5.5 μm. The basidia have basal clamps are mostly four-spored, and sometimes have cyanophilous granular contents.
Ramaria stricta has a cosmopolitan distribution, and is a fairly common species. The fungus is lignicolous, common in late summer and fall in coniferous forests of the Pacific Coast and Rocky Mountains. The fungus grows on dead wood, stumps, trunks, and branches of both leafy and coniferous trees. The form that grows on deciduous wood tends to be more orange and less bushy than those which grow on coniferous wood. Fruit bodies can form in "log lines" where decaying wood is buried underground near the surface, or is in an advanced state of decomposition.
Kingdom: Fungi
Division: Basidiomycota
Class: Agaricomycetes
Order: Gomphales
Family: Gomphaceae
Genus: Ramaria
Species: R. stricta
Binomial name Ramaria stricta
(Pers.) Quél. (1888)
Its fruit body is up to 10 cm (3.9 in) tall, made of multiple slender, compact, and vertical parallel branches. Its color is typically light tan to vinaceous-brown. All parts of the mushroom will bruise when handled.
There are several lookalike corals that can usually be distinguished from
R. stricta by differences in coloration, bruising reaction, or microscopic features. The fungus is inedible due to its unpleasant odor and bitter taste.
The species was originally described under the name Clavaria stricta by Christian Hendrik Persoon in 1795. In 1888, French mycologist Lucien Quélet transferred the species to the genus Ramaria. It is commonly known as the "upright coral".
The color of the fruit body is light yellow brown to paler toward extremities, the tips are light yellow; bruising light reddish brown. The basidiocarp has a leathery texture when fresh, but becomes brittle when dry. The stipe is branched up to 8 times, and the branches are all upright and nearly parallel. The branches ending in 4 to 5 thornlike tips. Overall, the fruit body appears bushy, and is medium-sized, up to 10 by 7 cm (3.9 by 2.8 in). The stipe is single or branching from the base; with white mycelium and rhizomorphs radiating from the base. The odor is of anise. The taste is bitter. The fruit bodies have been described as "edible but unpalatable," although other guides say the species is inedible.
The spore print is dark yellow. Spores are roughly elliptical, dotted with low cyanophilous warts, and measure 7–10 by 3.5–5.5 μm. The basidia have basal clamps are mostly four-spored, and sometimes have cyanophilous granular contents.
Ramaria stricta has a cosmopolitan distribution, and is a fairly common species. The fungus is lignicolous, common in late summer and fall in coniferous forests of the Pacific Coast and Rocky Mountains. The fungus grows on dead wood, stumps, trunks, and branches of both leafy and coniferous trees. The form that grows on deciduous wood tends to be more orange and less bushy than those which grow on coniferous wood. Fruit bodies can form in "log lines" where decaying wood is buried underground near the surface, or is in an advanced state of decomposition.
Kingdom: Fungi
Division: Basidiomycota
Class: Agaricomycetes
Order: Gomphales
Family: Gomphaceae
Genus: Ramaria
Species: R. stricta
Binomial name Ramaria stricta
(Pers.) Quél. (1888)

Mycena chlorophos
Bioluminescence Glow Mushroom
Bioluminescence Glow Mushroom


Mycena chlorophos is a species of agaric fungus in the family Mycenaceae. First described in 1860, the fungus is found in subtropical Asia, including Japan, Taiwan, Polynesia, Indonesia, and Sri Lanka, in Australia, and Brazil.
Mycena chlorophos the cap is initially convex before flattening out, fruit bodies (mushrooms) have pale brownish-grey sticky caps up to 30 mm (1.2 in) in diameter atop stems 6–30 mm (0.2–1.2 in) long and up to a millimeter thick. The cap has radial grooves extending to nearly the center, and sometimes develops cracks in the margin, which has small rounded teeth.
The white stem is 6–30 mm (0.24–1.18 in) long by 0.3–1 mm thick, hollow, and translucent. It has tiny hairs on the surface. The base of the stem is disc-shaped or somewhat bulbous, measuring 1–2.5 mm wide. The thin gills are free from attachment to the stem, or are adnexed (narrowly attached) to a slight collar encircling the stem. Initially white then grayish in color, they are somewhat crowded, with 17–32 full-length gills and 1 to 3 tiers of lamellulae (shorter gills that do not extend fully from the cap margin to the stem). The gills are 0.3–1 mm wide with micaceous edges. The flesh is very thin, and has a strong odor of ammonia.
M. chlorophos are bioluminescent and emit a pale green light. Both the caps and the gills are bioluminescent, while the mycelia and stems have little to no luminescence.
Fruiting occurs in forests on fallen woody debris such as dead twigs, branches, and logs. The fungus can be made to grow and fruit in laboratory conditions, and the growth conditions affecting bioluminescence have been investigated.
Fruit bodies of Mycena chlorophos are found in forests, where they grow in groups on woody debris such as fallen twigs, branches, and bark.
Bioluminescence studies :
Since the mushroom is small, and fruits in only a limited season in a small area, researchers have investigated the conditions needed to artificially cultivate the species in laboratory conditions, in order to have more material to study the mechanism of bioluminescence, and to help preserve the species.
The optimum temperature for the growth of mycelia is 27 °C (81 °F), while the optimum for the growth of primordia is 21 °C (70 °F). These temperatures are consistent with the subtropical climate in which the species is typically found. Maximum luminescence occurs at 27 °C, and about 25–39 hours after the primordia begin to form, when the cap has fully expanded. At 21 °C, luminescence persists for about 3 days, and becomes undetectable to the naked eyes about 72 hours after primordium initiation.
Class: Agaricomycetes
Order: Agaricales
Family: Mycenaceae
Genus: Mycena
Species: M. chlorophos
Binomial name Mycena chlorophos
(Berk. & M.A.Curtis) Sacc. (1887)
Mycena chlorophos the cap is initially convex before flattening out, fruit bodies (mushrooms) have pale brownish-grey sticky caps up to 30 mm (1.2 in) in diameter atop stems 6–30 mm (0.2–1.2 in) long and up to a millimeter thick. The cap has radial grooves extending to nearly the center, and sometimes develops cracks in the margin, which has small rounded teeth.
The white stem is 6–30 mm (0.24–1.18 in) long by 0.3–1 mm thick, hollow, and translucent. It has tiny hairs on the surface. The base of the stem is disc-shaped or somewhat bulbous, measuring 1–2.5 mm wide. The thin gills are free from attachment to the stem, or are adnexed (narrowly attached) to a slight collar encircling the stem. Initially white then grayish in color, they are somewhat crowded, with 17–32 full-length gills and 1 to 3 tiers of lamellulae (shorter gills that do not extend fully from the cap margin to the stem). The gills are 0.3–1 mm wide with micaceous edges. The flesh is very thin, and has a strong odor of ammonia.
M. chlorophos are bioluminescent and emit a pale green light. Both the caps and the gills are bioluminescent, while the mycelia and stems have little to no luminescence.
Fruiting occurs in forests on fallen woody debris such as dead twigs, branches, and logs. The fungus can be made to grow and fruit in laboratory conditions, and the growth conditions affecting bioluminescence have been investigated.
Fruit bodies of Mycena chlorophos are found in forests, where they grow in groups on woody debris such as fallen twigs, branches, and bark.
Bioluminescence studies :
Since the mushroom is small, and fruits in only a limited season in a small area, researchers have investigated the conditions needed to artificially cultivate the species in laboratory conditions, in order to have more material to study the mechanism of bioluminescence, and to help preserve the species.
The optimum temperature for the growth of mycelia is 27 °C (81 °F), while the optimum for the growth of primordia is 21 °C (70 °F). These temperatures are consistent with the subtropical climate in which the species is typically found. Maximum luminescence occurs at 27 °C, and about 25–39 hours after the primordia begin to form, when the cap has fully expanded. At 21 °C, luminescence persists for about 3 days, and becomes undetectable to the naked eyes about 72 hours after primordium initiation.
Class: Agaricomycetes
Order: Agaricales
Family: Mycenaceae
Genus: Mycena
Species: M. chlorophos
Binomial name Mycena chlorophos
(Berk. & M.A.Curtis) Sacc. (1887)
How is bioluminescence form
Bioluminescence is the production and emission of light by a living organism. It is a form of chemiluminescence.
Bioluminescence occurs widely in marine vertebrates and invertebrates, as well as in some fungi, microorganisms including some bioluminescent bacteria, and terrestrial arthropods such as fireflies.
Bioluminescence is used by living things to hunt prey, defend against predators, find mates, and execute other vital activities and some species luminesce to confuse attackers.
Most bioluminescent organisms are found in the ocean. These bioluminescent marine species include fish, bacteria, and jellies and Some bioluminescent organisms, including fireflies and fungi, are found on land. There are almost no bioluminescent organisms native to freshwater habitats.
In some animals, the light is bacteriogenic, produced by symbiotic bacteria such as those from the genus Vibrio; in others, it is autogenic, produced by the animals themselves.
Bioluminescence is a "cold light." Cold light means less than 20% of the light generates thermal radiation, or heat.
Luciferin is the compound that actually produces light. Luciferase is an enzyme. An enzyme is a chemical (called a catalyst) that interacts with a substrate to affect the rate of a chemical reaction.
In a general sense, the principal chemical reaction in bioluminescence involves a light-emitting molecule and an enzyme, generally called luciferin and luciferase, respectively.
In a chemical reaction, luciferin is called the substrate. The bioluminescent color (yellow in fireflies, greenish in lanternfish) is a result of the arrangement of luciferin molecules.
Some bioluminescent organisms produce (synthesize) luciferin on their own and some bioluminescent organisms do not synthesize luciferin. Instead, they absorb it through other organisms, either as food or in a symbiotic relationship.
The interaction of the luciferase with oxidized (oxygen-added) luciferin creates a byproduct, called oxyluciferin which is a type of chemical reaction creates light.
Most bioluminescent reactions involve luciferin and luciferase. Some reactions, however, do not involve an enzyme (luciferase). These reactions involve a chemical called a photoprotein. Photoproteins combine with luciferins and oxygen, but need another agent, often an ion of the element calcium, to produce light.
The appearance of bioluminescent light varies greatly, depending on the habitat and organism in which it is found.
Some species of fungi present in decaying wood, for instance, emit a fairly consistent glow, called foxfire and some organisms emit very bright light continuously.
Bioluminescence occurs widely in marine vertebrates and invertebrates, as well as in some fungi, microorganisms including some bioluminescent bacteria, and terrestrial arthropods such as fireflies.
Bioluminescence is used by living things to hunt prey, defend against predators, find mates, and execute other vital activities and some species luminesce to confuse attackers.
Most bioluminescent organisms are found in the ocean. These bioluminescent marine species include fish, bacteria, and jellies and Some bioluminescent organisms, including fireflies and fungi, are found on land. There are almost no bioluminescent organisms native to freshwater habitats.
In some animals, the light is bacteriogenic, produced by symbiotic bacteria such as those from the genus Vibrio; in others, it is autogenic, produced by the animals themselves.
Bioluminescence is a "cold light." Cold light means less than 20% of the light generates thermal radiation, or heat.
Luciferin is the compound that actually produces light. Luciferase is an enzyme. An enzyme is a chemical (called a catalyst) that interacts with a substrate to affect the rate of a chemical reaction.
In a general sense, the principal chemical reaction in bioluminescence involves a light-emitting molecule and an enzyme, generally called luciferin and luciferase, respectively.
In a chemical reaction, luciferin is called the substrate. The bioluminescent color (yellow in fireflies, greenish in lanternfish) is a result of the arrangement of luciferin molecules.
Some bioluminescent organisms produce (synthesize) luciferin on their own and some bioluminescent organisms do not synthesize luciferin. Instead, they absorb it through other organisms, either as food or in a symbiotic relationship.
The interaction of the luciferase with oxidized (oxygen-added) luciferin creates a byproduct, called oxyluciferin which is a type of chemical reaction creates light.
Most bioluminescent reactions involve luciferin and luciferase. Some reactions, however, do not involve an enzyme (luciferase). These reactions involve a chemical called a photoprotein. Photoproteins combine with luciferins and oxygen, but need another agent, often an ion of the element calcium, to produce light.
The appearance of bioluminescent light varies greatly, depending on the habitat and organism in which it is found.
Some species of fungi present in decaying wood, for instance, emit a fairly consistent glow, called foxfire and some organisms emit very bright light continuously.
Why Do Some Mushrooms Glow In The Dark?
Why Do Some Mushrooms Glow In The Dark? Glowing in the dark costs energy.
The Secret Behind Bioluminescent Mushrooms’ Magic Glow.
Scientists use chemistry to account for an astonishing phenomenon. Researchers described the compound that gives the mushrooms their glow — It’s called oxyluciferin.
Mushrooms use luciferins—light-emitting compounds found in other glowing animals and plants—to attract insects. The bugs then help spread their spores to sheltered places in the forest, which helps the mushroom species survive.
Luciferins give fireflies and even bioluminescent underwater creatures their glow. Paired with an enzyme and oxygen, it releases light that illuminates the fungi.
Scientists went foraging for the glow-in-the-dark mushrooms and they crushed the mushrooms to make a slurry filled with luciferins. Then they isolated the luciferin and studied it, capturing its chemical structure and experimenting with its ability to fuel those flourescent colors.
Not only does the team now know that the mushrooms are fueled by their own kind of luciferin, but they also figured out that the enzyme that combines with the chemical to trigger light could be what they call “promiscuous.”
That means that the enzyme might be able to interact with different luciferins—and produce even more shades of that pretty glow. And that suggests that when it comes to these magical mushrooms, there’s even more to discover.
Fungi produce tiny spores to spread themselves, much like seeds from trees. Insects can help transport the fungal spores. Fruit bodies are produced to disperse their sexual progeny as spores. Many fungi shoot spores into the air from the underside of the mushrooms, relying on moving air currents to passively distribute the spores over a wide area.
Through aroma. Truffles, the fruiting body of the Ascomycete fungi, use their smell to attract fungivores such as pigs or squirrels who eat them and leave spores behind in their waste. Stinkhorn mushrooms have a foul-smelling slime which attracts flies and other insects. The flies eat the slime and unwittingly spread the spores elsewhere.
Since bioluminescence is so energetically costly, the mushrooms only glow at night. They don’t glow bright enough during the day to be extra visible. An added bonus is that spores prefer to become active and grow at night when it is more humid.
Bioluminescence is one of the factors attracting insects to the mushrooms to help them spread their spores.
"On a totally dark night, without any moon, if you have your light off, these green mushrooms are basically the only light source you see in the forest besides the fireflies."
You just have to turn off your flashlight and the mushrooms stand out if they're there.
The Secret Behind Bioluminescent Mushrooms’ Magic Glow.
Scientists use chemistry to account for an astonishing phenomenon. Researchers described the compound that gives the mushrooms their glow — It’s called oxyluciferin.
Mushrooms use luciferins—light-emitting compounds found in other glowing animals and plants—to attract insects. The bugs then help spread their spores to sheltered places in the forest, which helps the mushroom species survive.
Luciferins give fireflies and even bioluminescent underwater creatures their glow. Paired with an enzyme and oxygen, it releases light that illuminates the fungi.
Scientists went foraging for the glow-in-the-dark mushrooms and they crushed the mushrooms to make a slurry filled with luciferins. Then they isolated the luciferin and studied it, capturing its chemical structure and experimenting with its ability to fuel those flourescent colors.
Not only does the team now know that the mushrooms are fueled by their own kind of luciferin, but they also figured out that the enzyme that combines with the chemical to trigger light could be what they call “promiscuous.”
That means that the enzyme might be able to interact with different luciferins—and produce even more shades of that pretty glow. And that suggests that when it comes to these magical mushrooms, there’s even more to discover.
Fungi produce tiny spores to spread themselves, much like seeds from trees. Insects can help transport the fungal spores. Fruit bodies are produced to disperse their sexual progeny as spores. Many fungi shoot spores into the air from the underside of the mushrooms, relying on moving air currents to passively distribute the spores over a wide area.
Through aroma. Truffles, the fruiting body of the Ascomycete fungi, use their smell to attract fungivores such as pigs or squirrels who eat them and leave spores behind in their waste. Stinkhorn mushrooms have a foul-smelling slime which attracts flies and other insects. The flies eat the slime and unwittingly spread the spores elsewhere.
Since bioluminescence is so energetically costly, the mushrooms only glow at night. They don’t glow bright enough during the day to be extra visible. An added bonus is that spores prefer to become active and grow at night when it is more humid.
Bioluminescence is one of the factors attracting insects to the mushrooms to help them spread their spores.
"On a totally dark night, without any moon, if you have your light off, these green mushrooms are basically the only light source you see in the forest besides the fireflies."
You just have to turn off your flashlight and the mushrooms stand out if they're there.
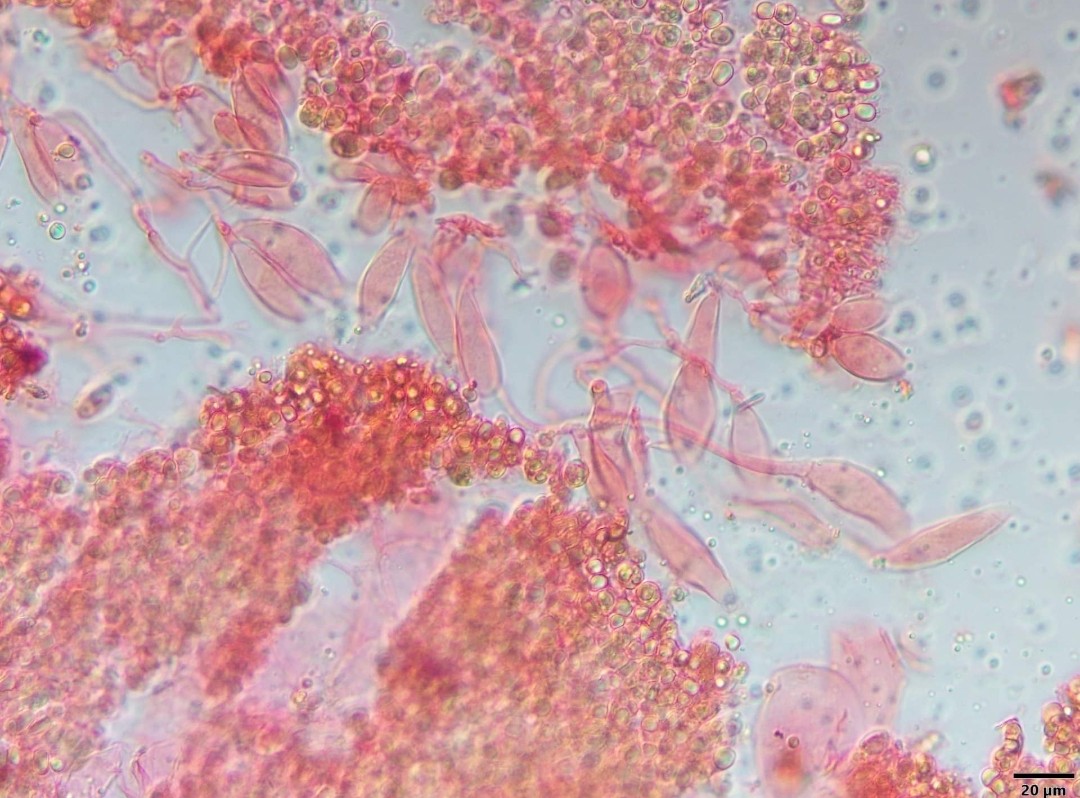
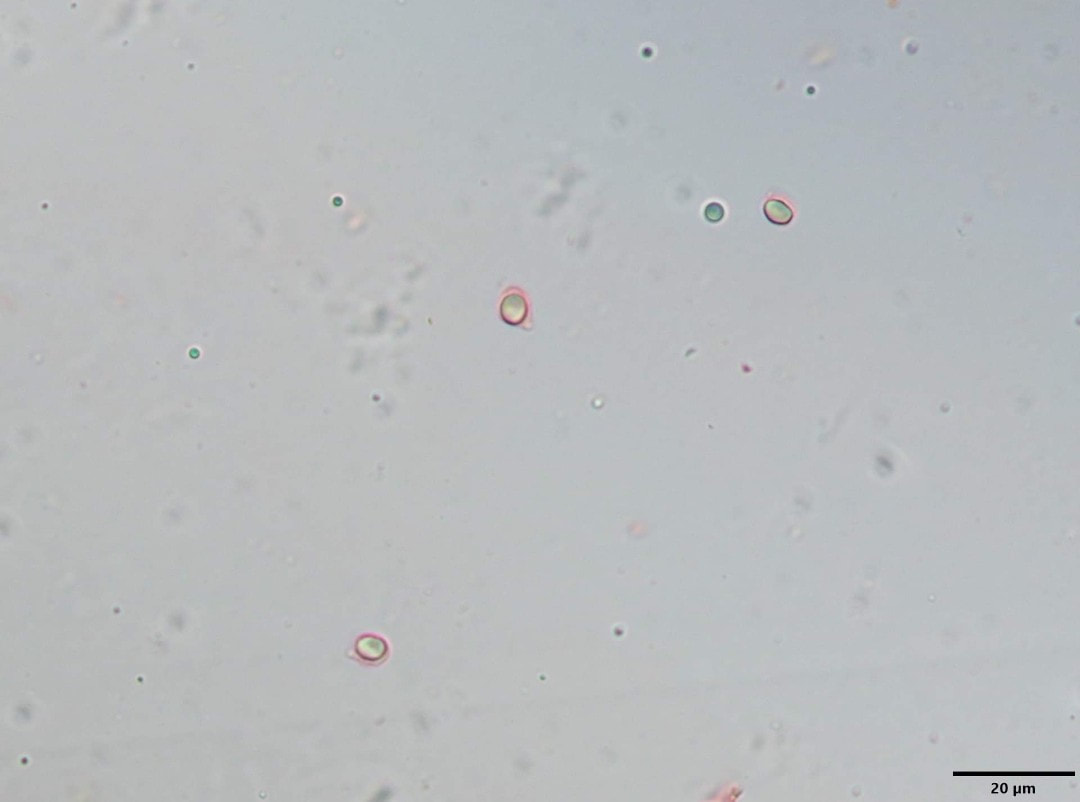
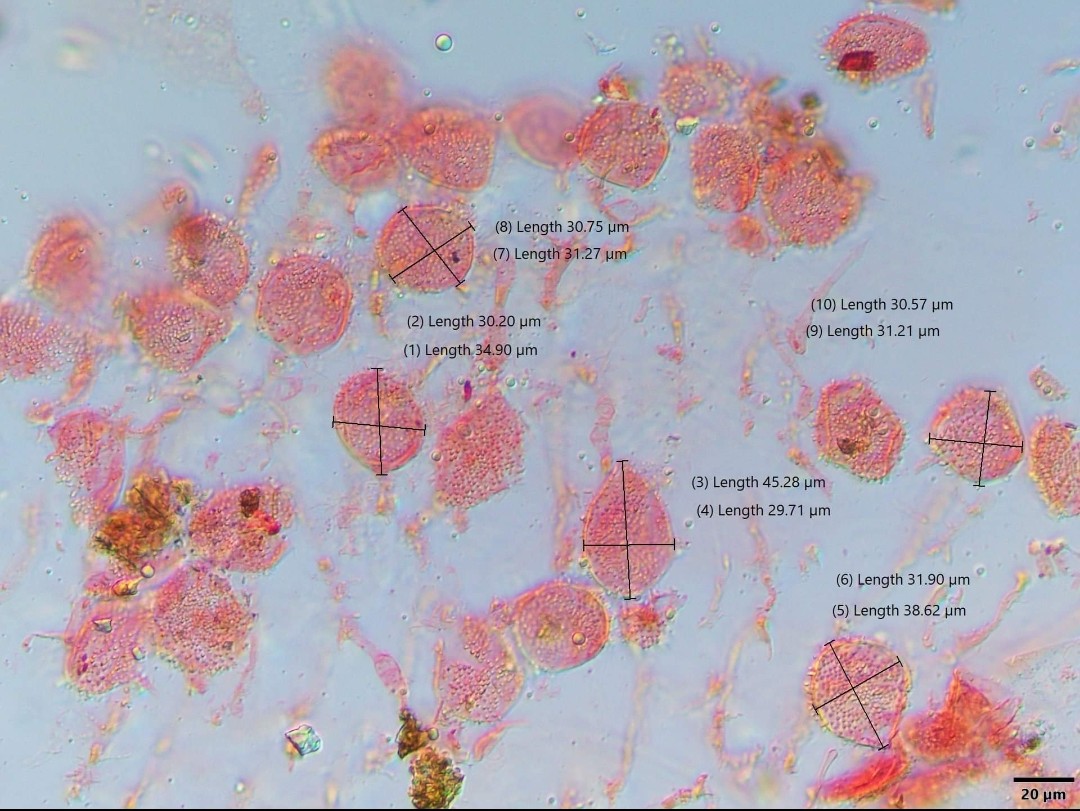











Lap test species confimation on 19 Dec 2020
Credit : Serena Lee
Credit : Serena Lee





Auricularia auricula-judae 黑木耳
The Jew's ear, (black) wood ear
The Jew's ear, (black) wood ear

Auricularia auricula-judae (Latin for 'Judas's Ear'), known as the Jew's ear, (black) wood ear, jelly ear, pepeao or by a number of other common names, is a species of edible Auriculariales fungus found worldwide.
The fruiting body is distinguished by its noticeably ear-like shape and brown colouration; it grows upon wood, especially elder. The fungus can be found throughout the year in temperate regions worldwide, where it grows upon both dead and living wood.
In the West, A. auricula-judae was used in folk medicine as recently as the 19th century for complaints including sore throats, sore eyes and jaundice, and as an astringent. Although it is not widely consumed in the West, it has long been popular in China. Today, the fungus is a popular ingredient in many Chinese dishes, such as hot and sour soup, and also used in Chinese medicine. It is also used in Ghana, as a blood tonic. Modern research into possible medical applications has variously concluded that A. auricula-judae has antitumour, hypoglycemic, anticoagulant and cholesterol-lowering properties.
The fruit body of A. auricula-judae is normally 3 to 8 centimetres (1.2 to 3.1 in) across, but can be as much as 12 centimetres (4.7 in). It is distinctively shaped, typically being reminiscent of a floppy ear, though the fruit bodies can also be cup-shaped. It is normally attached to the substrate laterally and sometimes by a very short stalk. The species has a tough, gelatinous, elastic texture when fresh, but it dries hard and brittle. The outer surface is a bright reddish-tan-brown with a purplish hint, often covered in tiny, downy hairs of a grey colour. It can be smooth, as is typical of younger specimens, or undulating with folds and wrinkles. The colour becomes darker with age. The inner surface is a lighter grey-brown in colour and smooth. It is sometimes wrinkled, again with folds and wrinkles, and may have "veins", making it appear even more ear-like.
The spores of A. auricula-judae are long and sausage shaped, ranging in size from 16 to 18 micrometres (μm) long by 6 to 8 μm thick. The spores themselves are white, cream or yellowish, and are hyaline. The spores can sometimes be seen in a whitish mass on the underside of the fruit body. The species has elongated cylindrical basidia with three transverse septa (internal cross-walls dividing the hyphae). Basidia 60-72x 4–7.5μm; sterigmata lateral, well developed, 3–4.5μm long. Spores smooth, hyaline, reniform to allantoid, 14-18 Χ 6-8μm, guttulate. Hairs on the fruit body are from 85 to 100 μm in length, and 5 to 6 μm in diameter. They are hyaline, lack a central strand and have rounded tips. They do not grow in dense tufts.
A. auricula-judae are popular in China, and are known by the general name of "wood ears" (木耳).
The medicinal use of food is common in China: a soup containing the species is used medicinally for dealing with colds and fevers by reducing the heat of the body.
Class: Agaricomycetes
Order: Auriculariales
Family: Auriculariaceae
Genus: Auricularia
Species: A. auricula-judae
The fruiting body is distinguished by its noticeably ear-like shape and brown colouration; it grows upon wood, especially elder. The fungus can be found throughout the year in temperate regions worldwide, where it grows upon both dead and living wood.
In the West, A. auricula-judae was used in folk medicine as recently as the 19th century for complaints including sore throats, sore eyes and jaundice, and as an astringent. Although it is not widely consumed in the West, it has long been popular in China. Today, the fungus is a popular ingredient in many Chinese dishes, such as hot and sour soup, and also used in Chinese medicine. It is also used in Ghana, as a blood tonic. Modern research into possible medical applications has variously concluded that A. auricula-judae has antitumour, hypoglycemic, anticoagulant and cholesterol-lowering properties.
The fruit body of A. auricula-judae is normally 3 to 8 centimetres (1.2 to 3.1 in) across, but can be as much as 12 centimetres (4.7 in). It is distinctively shaped, typically being reminiscent of a floppy ear, though the fruit bodies can also be cup-shaped. It is normally attached to the substrate laterally and sometimes by a very short stalk. The species has a tough, gelatinous, elastic texture when fresh, but it dries hard and brittle. The outer surface is a bright reddish-tan-brown with a purplish hint, often covered in tiny, downy hairs of a grey colour. It can be smooth, as is typical of younger specimens, or undulating with folds and wrinkles. The colour becomes darker with age. The inner surface is a lighter grey-brown in colour and smooth. It is sometimes wrinkled, again with folds and wrinkles, and may have "veins", making it appear even more ear-like.
The spores of A. auricula-judae are long and sausage shaped, ranging in size from 16 to 18 micrometres (μm) long by 6 to 8 μm thick. The spores themselves are white, cream or yellowish, and are hyaline. The spores can sometimes be seen in a whitish mass on the underside of the fruit body. The species has elongated cylindrical basidia with three transverse septa (internal cross-walls dividing the hyphae). Basidia 60-72x 4–7.5μm; sterigmata lateral, well developed, 3–4.5μm long. Spores smooth, hyaline, reniform to allantoid, 14-18 Χ 6-8μm, guttulate. Hairs on the fruit body are from 85 to 100 μm in length, and 5 to 6 μm in diameter. They are hyaline, lack a central strand and have rounded tips. They do not grow in dense tufts.
A. auricula-judae are popular in China, and are known by the general name of "wood ears" (木耳).
The medicinal use of food is common in China: a soup containing the species is used medicinally for dealing with colds and fevers by reducing the heat of the body.
Class: Agaricomycetes
Order: Auriculariales
Family: Auriculariaceae
Genus: Auricularia
Species: A. auricula-judae


Tremella fuciformis 白木耳

Tremella fuciformis is a species of fungus; it produces white, frond-like, gelatinous basidiocarps (fruiting bodies). It is widespread, especially in the tropics, where it can be found on the dead branches of broadleaf trees. This fungus is commercially cultivated and is one of the most popular fungi in the cuisine and medicine of China.
Tremella fuciformis is commonly known as snow fungus, snow ear, silver ear fungus, and white jelly mushroom.
In Mandarin Chinese, it is called 银耳 (pinyin: "silver ear ), 雪耳 (pinyin: xuě ěr; snow ear ); or 白木耳 (pinyin: bái mù ěr, white wood ear ), and in Japanese it is called shiro kikurage (シロキクラゲ, lit. "white tree jellyfish"). In Vietnam, it is called nấm tuyết or ngân nhĩ.
Tremella fuciformis is known to be a parasite of Hypoxylon species, fruit bodies are typically found on dead, attached or recently fallen branches of broadleaf trees.
Tremella fuciformis has been cultivated in China since at least the nineteenth century. Initially, suitable wooden poles were prepared and then treated in various ways in the hope that they would be colonized by the fungus. This haphazard method of cultivation was improved when poles were inoculated with spores or mycelium. Modern production only began, however, with the realization that both the Tremella and its host species needed to be inoculated into the substrate to ensure success. The "dual culture" method, now used commercially, employs a sawdust mix inoculated with both fungal species and kept under optimal conditions.
Class: Tremellomycetes
Order: Tremellales
Family: Tremellaceae
Genus: Tremella
Species: T. fuciformis
Binomial name Tremella fuciformis
Tremella fuciformis is commonly known as snow fungus, snow ear, silver ear fungus, and white jelly mushroom.
In Mandarin Chinese, it is called 银耳 (pinyin: "silver ear ), 雪耳 (pinyin: xuě ěr; snow ear ); or 白木耳 (pinyin: bái mù ěr, white wood ear ), and in Japanese it is called shiro kikurage (シロキクラゲ, lit. "white tree jellyfish"). In Vietnam, it is called nấm tuyết or ngân nhĩ.
Tremella fuciformis is known to be a parasite of Hypoxylon species, fruit bodies are typically found on dead, attached or recently fallen branches of broadleaf trees.
Tremella fuciformis has been cultivated in China since at least the nineteenth century. Initially, suitable wooden poles were prepared and then treated in various ways in the hope that they would be colonized by the fungus. This haphazard method of cultivation was improved when poles were inoculated with spores or mycelium. Modern production only began, however, with the realization that both the Tremella and its host species needed to be inoculated into the substrate to ensure success. The "dual culture" method, now used commercially, employs a sawdust mix inoculated with both fungal species and kept under optimal conditions.
Class: Tremellomycetes
Order: Tremellales
Family: Tremellaceae
Genus: Tremella
Species: T. fuciformis
Binomial name Tremella fuciformis


Schizophyllum commune
split-gill mushroom
split-gill mushroom

Schizophyllum commune is a species of fungus in the genus Schizophyllum. The mushroom resembles undulating waves of tightly packed corals or loose Chinese fan. ”Gillies” or Split Gills vary from creamy yellow to pale white in colour. The cap is small, 1–4.5 cm wide with a dense yet spongey body texture. It is known as the split-gill mushroom because of the unique longitudinally divided nature of the "gills" on the underside of the cap. It is the only known fungi capable of retracting by movement. This mushroom is found throughout the world.
It is found in the wild on decaying trees after rainy seasons followed by dry spells where the mushrooms are naturally collected. It is known for its high medicinal value and aromatic taste profile. It has recently attracted the medicinal industry for its immunomodulatory, antifungal, antineoplastic and antiviral activities that are higher than those of any other glucan complex carbohydrate.
S. commune is usually described as a morphological species of global distribution, but some research has suggested that it may be a species complexencompassing several cryptic species of more narrow distribution, as typical of many mushroom-forming Basidiomycota.
The gills, which produce basidiospores on their surface, split when the mushroom dries out, earning this mushroom the common name split gill. It is common in rotting wood, but can also cause disease in humans.
It has 23,328 distinct sexes, properly called mating types. Individuals of any sex are compatible for mating with all but their own sex. However, there are two genetic loci determining the mating type, locus A with 288 alleles and locus B with 81 alleles. A pair of fungi will only be fertile if they have different A and different B alleles; that is, each sex can enter fertile pairings with 22,960 others.
Although European and US guidebooks list it as inedible, this is apparently due to differing standards of taste rather than known toxicity, being regarded with little culinary interest due to its tough texture. S. commune is, in fact, edible and widely consumed in Mexico and elsewhere in the tropics. In Northeast India, in the state Manipur it is known as kanglayen and one of the favourite ingredients for Manipuri-style pancakes called paaknam. In Mizoram, the local name is pasi (pa means mushroom, simeans tiny) and it is one of the highest rated edible mushrooms among the Mizo community. The authors explain the preference for tough, rubbery mushrooms in the tropics as a consequence of the fact that tender, fleshy mushrooms quickly rot in the hot humid conditions there, making their marketing problematic.
Class: Agaricomycetes
Order: Agaricales
Family: Schizophyllaceae
Genus: Schizophyllum
Species: S. commune
Binomial name Schizophyllum commune
Fries (1815)
It is found in the wild on decaying trees after rainy seasons followed by dry spells where the mushrooms are naturally collected. It is known for its high medicinal value and aromatic taste profile. It has recently attracted the medicinal industry for its immunomodulatory, antifungal, antineoplastic and antiviral activities that are higher than those of any other glucan complex carbohydrate.
S. commune is usually described as a morphological species of global distribution, but some research has suggested that it may be a species complexencompassing several cryptic species of more narrow distribution, as typical of many mushroom-forming Basidiomycota.
The gills, which produce basidiospores on their surface, split when the mushroom dries out, earning this mushroom the common name split gill. It is common in rotting wood, but can also cause disease in humans.
It has 23,328 distinct sexes, properly called mating types. Individuals of any sex are compatible for mating with all but their own sex. However, there are two genetic loci determining the mating type, locus A with 288 alleles and locus B with 81 alleles. A pair of fungi will only be fertile if they have different A and different B alleles; that is, each sex can enter fertile pairings with 22,960 others.
Although European and US guidebooks list it as inedible, this is apparently due to differing standards of taste rather than known toxicity, being regarded with little culinary interest due to its tough texture. S. commune is, in fact, edible and widely consumed in Mexico and elsewhere in the tropics. In Northeast India, in the state Manipur it is known as kanglayen and one of the favourite ingredients for Manipuri-style pancakes called paaknam. In Mizoram, the local name is pasi (pa means mushroom, simeans tiny) and it is one of the highest rated edible mushrooms among the Mizo community. The authors explain the preference for tough, rubbery mushrooms in the tropics as a consequence of the fact that tender, fleshy mushrooms quickly rot in the hot humid conditions there, making their marketing problematic.
Class: Agaricomycetes
Order: Agaricales
Family: Schizophyllaceae
Genus: Schizophyllum
Species: S. commune
Binomial name Schizophyllum commune
Fries (1815)

Ganoderma lucidum


Ganoderma lucidum is a reddish laccate species of Ganoderma with a limited distribution in Europe and parts of China, where it grows on decaying hardwood trees. Wild populations have been found in the United States in California and Utah, but were likely introduced anthropogenically and naturalized.
The history of the Ganoderma lucidum taxon is tied to the history of the erection of Ganoderma as a genus. Karsten first described the Ganoderma in 1881 and included only one species in the genus, G. lucidum (Curtis) Karst.
Previously, it was called Boletus lucidus Curtis (1781) and then Polyporus lucidus (Curtis) Fr. (1821). Patouillard revised Karsten’s genus Ganoderma to include all species with pigmented spores, adhering tubes and laccate crusted pilei, which resulted in a total of 48 species classified under the genus Ganoderma in his 1889 monograph.
Despite this recognition of additional species and subsequent discoveries of new Ganoderma species, such as 17 new North American species identified by Murrill North in 1902, the taxonomy of Ganoderma species has remained chaotic, and the species name Ganoderma lucidum continues to be used for most laccate Ganoderma species, including commonly misidentifying Ganoderma lingzhi (also known as reishi mushroom (Japan) or lingzhi/ling chih (China)), the sought-after red laccate Ganoderma species used in traditional Asian medicine.
It is important to note that G. lucidum is not a synonym for G. lingzhi and is not in the same clade: based on molecular phylogenetic analyses, G. lucidum is more closely related to North American species Ganoderma tsugae and Ganoderma oregonense than to G. lingzhi, whose sister taxa include Ganoderma curtisii and Ganoderma ravenelii.
These genetic analyses tested species concept hypotheses to determine how the laccate Ganoderma
taxa are related. One such study found six major clades among the 29 samples studied. Samples labeled as G. lucidum were found in five of the six clades, showing the extent of the confusion around species identification.
Another study found similar results, and also showed that Ganoderma resinaceum from Europe and the North American sample wrongly labeled G. lucidum were sister taxa and were also more closely related to each other than the European G. lucidum sensu stricto.
A recent multilocus phylogeny, using ITS, tef, rpb1, and rpb2, revealed that the global diversity of the laccate Ganoderma species included three supported major lineages. These results agree with several of the earlier works focusing mostly on morphology, geography and host preference, but with statistical support separating the European and North American taxa.
The phylogenetic species concept using a multilocus approach is currently the most robust and accepted method for designating species ranks for the fungi. The species is inedible but is used to make a bitter-tasting tea.
The confusion surrounding the taxonomy of Ganoderma species has nonetheless persisted, causing confusion and inaccuracies when labeling medicinal and manufactured products containing Ganoderma species, as well as "grow your own" (GYO) kits and other tissue samples sold for cultivation of Ganoderma species.
Products typically carry a label of G. lucidum, very often along with the words "reishi" and "lingzhi/ling chih" (which most typically refer to Asian Ganoderma species used in traditional medicine, such as G. lingzhi and Ganoderma sinense) merely because they contain a laccate Ganoderma species.
These products and GYO kits sold as Ganoderma lucidum very often do not contain G. lucidum sensu stricto: one study showed through DNA analysis that 93% of GYO kits and half of the dried mushroom products studied that were labeled "G. lucidum" contained G. lingzhi in actuality, a happy result for those wanting the Asian medicinal mushroom but inaccurate labeling nonetheless.
The study also found that no manufactured reishi product and only one GYO kit actually contained G. lucidum sensu stricto. Other species present in these products included Ganoderma applanatum, Ganoderma australe (potentially a species complex), Ganoderma gibbosum, Ganoderma sessile, and G. sinense.
Ganoderma lucidum sensu stricto has a limited distribution in Europe and parts of China, where it grows on the decaying hardwood trees.
The fruiting body almost always has a stipe present, which is tawny to russet colored and 1.5 times the diameter of the cap. Context tissue (sterile tissue inside the fruiting body between the pileus crust and the initiation of the tubes) is pink-buff to cinnamon-buff and corky, showing concentric growth zones and no resinous or melanoid deposits. The hymenium displays 4-5 pores per millimetre. Chlamydospores are absent. Basidiospores are 8.2-12.1μm (average 10.7) long, 4.8-8.9μm (average 7.1) wide, a spore shape index (SSI) of 66.2 and a Q ratio of 1.5.
Division: Basidiomycota
Class: Agaricomycetes
Order: Polyporales
Family: Ganodermataceae
Genus: Ganoderma
Species: G. lucidum
Binomial name Ganoderma lucidum
Karst (1881)
The history of the Ganoderma lucidum taxon is tied to the history of the erection of Ganoderma as a genus. Karsten first described the Ganoderma in 1881 and included only one species in the genus, G. lucidum (Curtis) Karst.
Previously, it was called Boletus lucidus Curtis (1781) and then Polyporus lucidus (Curtis) Fr. (1821). Patouillard revised Karsten’s genus Ganoderma to include all species with pigmented spores, adhering tubes and laccate crusted pilei, which resulted in a total of 48 species classified under the genus Ganoderma in his 1889 monograph.
Despite this recognition of additional species and subsequent discoveries of new Ganoderma species, such as 17 new North American species identified by Murrill North in 1902, the taxonomy of Ganoderma species has remained chaotic, and the species name Ganoderma lucidum continues to be used for most laccate Ganoderma species, including commonly misidentifying Ganoderma lingzhi (also known as reishi mushroom (Japan) or lingzhi/ling chih (China)), the sought-after red laccate Ganoderma species used in traditional Asian medicine.
It is important to note that G. lucidum is not a synonym for G. lingzhi and is not in the same clade: based on molecular phylogenetic analyses, G. lucidum is more closely related to North American species Ganoderma tsugae and Ganoderma oregonense than to G. lingzhi, whose sister taxa include Ganoderma curtisii and Ganoderma ravenelii.
These genetic analyses tested species concept hypotheses to determine how the laccate Ganoderma
taxa are related. One such study found six major clades among the 29 samples studied. Samples labeled as G. lucidum were found in five of the six clades, showing the extent of the confusion around species identification.
Another study found similar results, and also showed that Ganoderma resinaceum from Europe and the North American sample wrongly labeled G. lucidum were sister taxa and were also more closely related to each other than the European G. lucidum sensu stricto.
A recent multilocus phylogeny, using ITS, tef, rpb1, and rpb2, revealed that the global diversity of the laccate Ganoderma species included three supported major lineages. These results agree with several of the earlier works focusing mostly on morphology, geography and host preference, but with statistical support separating the European and North American taxa.
The phylogenetic species concept using a multilocus approach is currently the most robust and accepted method for designating species ranks for the fungi. The species is inedible but is used to make a bitter-tasting tea.
The confusion surrounding the taxonomy of Ganoderma species has nonetheless persisted, causing confusion and inaccuracies when labeling medicinal and manufactured products containing Ganoderma species, as well as "grow your own" (GYO) kits and other tissue samples sold for cultivation of Ganoderma species.
Products typically carry a label of G. lucidum, very often along with the words "reishi" and "lingzhi/ling chih" (which most typically refer to Asian Ganoderma species used in traditional medicine, such as G. lingzhi and Ganoderma sinense) merely because they contain a laccate Ganoderma species.
These products and GYO kits sold as Ganoderma lucidum very often do not contain G. lucidum sensu stricto: one study showed through DNA analysis that 93% of GYO kits and half of the dried mushroom products studied that were labeled "G. lucidum" contained G. lingzhi in actuality, a happy result for those wanting the Asian medicinal mushroom but inaccurate labeling nonetheless.
The study also found that no manufactured reishi product and only one GYO kit actually contained G. lucidum sensu stricto. Other species present in these products included Ganoderma applanatum, Ganoderma australe (potentially a species complex), Ganoderma gibbosum, Ganoderma sessile, and G. sinense.
Ganoderma lucidum sensu stricto has a limited distribution in Europe and parts of China, where it grows on the decaying hardwood trees.
The fruiting body almost always has a stipe present, which is tawny to russet colored and 1.5 times the diameter of the cap. Context tissue (sterile tissue inside the fruiting body between the pileus crust and the initiation of the tubes) is pink-buff to cinnamon-buff and corky, showing concentric growth zones and no resinous or melanoid deposits. The hymenium displays 4-5 pores per millimetre. Chlamydospores are absent. Basidiospores are 8.2-12.1μm (average 10.7) long, 4.8-8.9μm (average 7.1) wide, a spore shape index (SSI) of 66.2 and a Q ratio of 1.5.
Division: Basidiomycota
Class: Agaricomycetes
Order: Polyporales
Family: Ganodermataceae
Genus: Ganoderma
Species: G. lucidum
Binomial name Ganoderma lucidum
Karst (1881)
**********************************
Lingzhi, Ganoderma lingzhi, also known as reishi, is a polypore fungus ("bracket fungus") belonging to the genus Ganoderma.
Its red-varnished, kidney-shaped cap and peripherally inserted stem gives it a distinct fan-like appearance. When fresh, the lingzhi is soft, cork-like, and flat. It lacks gills on its underside, and instead releases its spores via fine pores. Depending on the age, the pores on its underside may be white or brown.
The lingzhi mushroom is used in traditional Chinese medicine, but there is insufficient evidence that it is effective for treating any disease. In nature, it grows at the base and stumps of deciduous trees, especially that of the maple. Only two or three out of 10,000 such aged trees will have lingzhi growth, and therefore its wild form is rare. Lingzhi may be cultivated on hardwood logs, sawdust or woodchips.
Ganoderma lingzhi is found in Asia growing as a parasite or saprotroph on a variety of trees.
Ganoderma curtisii and Ganoderma ravenelii are the closest relatives of the lingzhi mushroom in North America.
In the wild, lingzhi grows at the base and stumps of deciduous trees, especially that of the maple. Only two or three out of 10,000 such aged trees will have lingzhi growth, and therefore it is extremely rare in its natural form. Today, lingzhi is effectively cultivated on hardwood logs or sawdust/woodchips.
Because of its bitter taste, lingzhi is traditionally prepared as a hot water extract product. Thinly sliced or pulverized lingzhi (either fresh or dried) is added to boiling water which is then reduced to a simmer, covered, and left for 2 hours. The resulting liquid is dark and fairly bitter in taste. The red lingzhi is often more bitter than the black. The process is sometimes repeated to increase the concentration. Alternatively, it can be used as an ingredient in a formula decoction, or used to make an extract (in liquid, capsule, or powder form).
Lingzhi is now commercially manufactured and sold. Since the early 1970s, most lingzhi is cultivated. Lingzhi can grow on substrates such as sawdust, grain, and wood logs. After formation of the fruiting body, lingzhi is most commonly harvested, dried, ground, and processed into tablets or capsules to be directly ingested or made into tea or soup. Other lingzhi products include processed fungal mycelia or spores.
Order: Polyporales
Family: Ganodermataceae
Genus: Ganoderma
Species: G. lingzhi
Binomial name Ganoderma lingzhi
(Sheng H. Wu, Y. Cao & Y.C. Dai) (2012)
Its red-varnished, kidney-shaped cap and peripherally inserted stem gives it a distinct fan-like appearance. When fresh, the lingzhi is soft, cork-like, and flat. It lacks gills on its underside, and instead releases its spores via fine pores. Depending on the age, the pores on its underside may be white or brown.
The lingzhi mushroom is used in traditional Chinese medicine, but there is insufficient evidence that it is effective for treating any disease. In nature, it grows at the base and stumps of deciduous trees, especially that of the maple. Only two or three out of 10,000 such aged trees will have lingzhi growth, and therefore its wild form is rare. Lingzhi may be cultivated on hardwood logs, sawdust or woodchips.
Ganoderma lingzhi is found in Asia growing as a parasite or saprotroph on a variety of trees.
Ganoderma curtisii and Ganoderma ravenelii are the closest relatives of the lingzhi mushroom in North America.
In the wild, lingzhi grows at the base and stumps of deciduous trees, especially that of the maple. Only two or three out of 10,000 such aged trees will have lingzhi growth, and therefore it is extremely rare in its natural form. Today, lingzhi is effectively cultivated on hardwood logs or sawdust/woodchips.
Because of its bitter taste, lingzhi is traditionally prepared as a hot water extract product. Thinly sliced or pulverized lingzhi (either fresh or dried) is added to boiling water which is then reduced to a simmer, covered, and left for 2 hours. The resulting liquid is dark and fairly bitter in taste. The red lingzhi is often more bitter than the black. The process is sometimes repeated to increase the concentration. Alternatively, it can be used as an ingredient in a formula decoction, or used to make an extract (in liquid, capsule, or powder form).
Lingzhi is now commercially manufactured and sold. Since the early 1970s, most lingzhi is cultivated. Lingzhi can grow on substrates such as sawdust, grain, and wood logs. After formation of the fruiting body, lingzhi is most commonly harvested, dried, ground, and processed into tablets or capsules to be directly ingested or made into tea or soup. Other lingzhi products include processed fungal mycelia or spores.
Order: Polyporales
Family: Ganodermataceae
Genus: Ganoderma
Species: G. lingzhi
Binomial name Ganoderma lingzhi
(Sheng H. Wu, Y. Cao & Y.C. Dai) (2012)




Trametes versicolor
Turkey tail.
Turkey tail.

Trametes versicolor – also known as Coriolus versicolor and Polyporus versicolor – is a common polypore mushroom found throughout the world. Meaning 'of several colours', versicolor reliably describes this fungus that displays different colors. For example, because its shape and multiple colors are similar to those of a wild turkey, T. versicolor is commonly called turkey tail.
The top surface of the cap shows typical concentric zones of different colours. The flesh is 1–3 mm thick and has leathery texture. Older specimens, can have zones with green algae growing on them, thus appearing green. It commonly grows in tiled layers. The cap is rust-brown or darker brown, sometimes with blackish zones. The cap is flat, up to 8 × 5 x 0.5–1 cm in area. It is often triangular or round, with zones of fine hairs. The pore surface is whitish to light brown, pores round and with age twisted and labyrinthine. 3–8 pores per millimeter.
It may be eaten by caterpillars of the fungus moth Nemaxera betulinella and by maggots of the Platypezid fly Polyporivora picta. and the fungus gnat Mycetophila luctuosa, but is considered inedible to humans.
Trametes versicolor contains polysaccharides under basic research, including the protein-bound PSP and β-1,3 and β-1,4 glucans. The lipid fraction contains the lanostane-type tetracyclic triterpenoid sterol ergosta-7,22,dien-3β-ol as well as fungisterol and β-sitosterol.
This mushroom, known as yun zhi [雲芝] in China, is used in traditional Chinese medicine; it is supposed to boost the immune system and have anticarcinogenic properties. Its polysaccharides are thought to be the effective constituents.
Clinical trials in people with breast cancer, leukemias, and liver cancer remain inconclusive as of 2016.
Order: Polyporales
Family: Polyporaceae
Genus: Trametes
Species: T. versicolor
Binomial nameTrametes versicolor
(L.) Lloyd (1920)
云芝(Trametes versicolor,前称Coriolus versicolor或Polyporus versicolor),又称彩云革盖菌或瓦菌,是多孔菌科植物云芝的子实体或菌丝体。云芝源自中国的原始森林,于全国东南西北都有分布,寄生于海拔三千米以上的阔叶树和朽木上。 云芝是一种大型珍贵药用真菌,常用于抗癌治疗。
云芝体积较小,无柄,呈覆瓦状或莲座状。其菌盖较薄,约1-8cm宽,0.1-0.3cm厚,呈半圆形至贝壳状。革质表面有细长的绒毛,与多种颜色组成狭窄的同心环形,边沿较薄,呈波浪状。云芝菌肉为白色的。云芝外貌与云耳相近,但质地却很坚硬。
云芝一般生长在昏暗且湿度较高的地方,相对湿度约85 – 90%之间。由于菌丝适应力较强,可生长于3 - 31℃,而25 - 28℃为最佳;子实体则最适合生长于25 - 30℃。于温差较大的环境,可加快子实体的增长。另外,在通风良好的地方,菌丝能迅速生长,子实体能更健全地成长。云芝喜欢生长在较酸性的环境,约pH 4 – 5.6。
含有丰富的蛋白质、脂肪、多糖、多糖肽及所有葡聚糖、三萜类、木质素、多种氨基酸及微量元素。多糖肽中还另有五类单糖,包括鼠李糖、半乳糖、阿拉伯糖、甘露糖及木糖。
主治,
维基百科中的医疗相关内容仅供参考,详见医学声明。如需医疗服务或可靠意见,请咨询专业人士。
云芝味甘平淡,性微寒,归肝、脾、肺经。传统用法为健脾利湿、止咳平喘及刺激免疫。现时,除了用作增强免疫功能,含有的云芝多糖更可用于抗肿瘤,抑压不良细胞的繁殖。云芝能辅助顽疾的治疗,减少化疗、电疗所引起的脱发、呕吐、食欲不振、口腔溃烂等副作用,纾缓疗程时所带来的不适,增加病患者的存活率。而且,用于护肝、治疗肝脏问题的功效显著,更可改善呼吸系统。
The top surface of the cap shows typical concentric zones of different colours. The flesh is 1–3 mm thick and has leathery texture. Older specimens, can have zones with green algae growing on them, thus appearing green. It commonly grows in tiled layers. The cap is rust-brown or darker brown, sometimes with blackish zones. The cap is flat, up to 8 × 5 x 0.5–1 cm in area. It is often triangular or round, with zones of fine hairs. The pore surface is whitish to light brown, pores round and with age twisted and labyrinthine. 3–8 pores per millimeter.
It may be eaten by caterpillars of the fungus moth Nemaxera betulinella and by maggots of the Platypezid fly Polyporivora picta. and the fungus gnat Mycetophila luctuosa, but is considered inedible to humans.
Trametes versicolor contains polysaccharides under basic research, including the protein-bound PSP and β-1,3 and β-1,4 glucans. The lipid fraction contains the lanostane-type tetracyclic triterpenoid sterol ergosta-7,22,dien-3β-ol as well as fungisterol and β-sitosterol.
This mushroom, known as yun zhi [雲芝] in China, is used in traditional Chinese medicine; it is supposed to boost the immune system and have anticarcinogenic properties. Its polysaccharides are thought to be the effective constituents.
Clinical trials in people with breast cancer, leukemias, and liver cancer remain inconclusive as of 2016.
Order: Polyporales
Family: Polyporaceae
Genus: Trametes
Species: T. versicolor
Binomial nameTrametes versicolor
(L.) Lloyd (1920)
云芝(Trametes versicolor,前称Coriolus versicolor或Polyporus versicolor),又称彩云革盖菌或瓦菌,是多孔菌科植物云芝的子实体或菌丝体。云芝源自中国的原始森林,于全国东南西北都有分布,寄生于海拔三千米以上的阔叶树和朽木上。 云芝是一种大型珍贵药用真菌,常用于抗癌治疗。
云芝体积较小,无柄,呈覆瓦状或莲座状。其菌盖较薄,约1-8cm宽,0.1-0.3cm厚,呈半圆形至贝壳状。革质表面有细长的绒毛,与多种颜色组成狭窄的同心环形,边沿较薄,呈波浪状。云芝菌肉为白色的。云芝外貌与云耳相近,但质地却很坚硬。
云芝一般生长在昏暗且湿度较高的地方,相对湿度约85 – 90%之间。由于菌丝适应力较强,可生长于3 - 31℃,而25 - 28℃为最佳;子实体则最适合生长于25 - 30℃。于温差较大的环境,可加快子实体的增长。另外,在通风良好的地方,菌丝能迅速生长,子实体能更健全地成长。云芝喜欢生长在较酸性的环境,约pH 4 – 5.6。
含有丰富的蛋白质、脂肪、多糖、多糖肽及所有葡聚糖、三萜类、木质素、多种氨基酸及微量元素。多糖肽中还另有五类单糖,包括鼠李糖、半乳糖、阿拉伯糖、甘露糖及木糖。
主治,
维基百科中的医疗相关内容仅供参考,详见医学声明。如需医疗服务或可靠意见,请咨询专业人士。
云芝味甘平淡,性微寒,归肝、脾、肺经。传统用法为健脾利湿、止咳平喘及刺激免疫。现时,除了用作增强免疫功能,含有的云芝多糖更可用于抗肿瘤,抑压不良细胞的繁殖。云芝能辅助顽疾的治疗,减少化疗、电疗所引起的脱发、呕吐、食欲不振、口腔溃烂等副作用,纾缓疗程时所带来的不适,增加病患者的存活率。而且,用于护肝、治疗肝脏问题的功效显著,更可改善呼吸系统。

